Функциональная гистология коры больших полушарий
Кора головного мозга является высшим отделом центральной нервной системы. Она представляет собой тонкий слой нервной ткани, образующий множество складок. Общая площадь поверхности коры 2200 см 2 . Толщина коры колеблется от 1,3 до 4,5 мм. Объем коры равен примерно 600 см 3 . В состав коры головного мозга входит 10 9 – 10 10 нейронов и еще большее число глиальных клеток (рис. 2.9). В пределах коры наблюдается чередование слоев, содержащих преимущественно тела нервных клеток, со слоями, образованными в основном их аксонами, и поэтому на свежем срезе кора головного мозга выглядит полосатой. На основании формы и расположения нервных клеток в коре с типичным строением можно выделить шесть слоев; некоторые из них подразделяются на два или более вторичных слоев. На основании строения коры выделяют следующие основные зоны: новую кору (неокортекс), старую кору (архикортекс), древнюю кору (палеокортекс) и межуточную кору (периархикортикальную и перипалеокортикальную). Самая обширная зона коры – неокортекс. Неокортекс занимает дорсальную и латеральную поверхность больших полушарий, в то время как палеокортекс находится на базальной и медиальной поверхности полушарий.
Рис. 2.9. Клеточный состав и слои коры головного мозга
В неокортексе различают следующие слои:
I. Молекулярный слой (плексиформный). В этом слое имеется множество волокон, образующих густое тангенциальное поверхностное сплетение, однако в нем мало клеток. Здесь содержатся в основном звездообразные мелкие клетки, которые осуществляют местную интеграцию деятельности эфферентных нейронов.
II. Наружный зернистый слой. Содержит мелкие нейроны различной формы, которые имеют синаптические связи с нейронами молекулярного слоя на всем поперечнике коры. В его глубине располагаются малые пирамидные клетки.
III. Наружный пирамидный слой. Этот слой состоит из пирамидных клеток малой и средней величины. Некоторые отделы коры в этом слое содержат крупные пирамидные клетки. Особенно много крупных пирамидных клеток в коре передней центральной извилины. Часть отростков этих клеток достигает первого слоя, участвуя в формировании тангенциального подслоя, другие погружаются в белое вещество полушарий мозга, поэтому III слой иногда обозначают как третичный ассоциативный.
IV. Внутренний зернистый слой. Характеризуется рыхлым расположением мелких нейронов различной величины и формы с преобладанием звездчатых, имеющих дугообразные возвратные аксоны. Аксоны клеток проникают в выше- и нижележащие слои. Звездчатые клетки представляют систему переключений с афферентных на эфферентные нейроны III и IV слоев. В IV слое также образуется тангенциальная прослойка из нервных волокон. Поэтому иногда этот слой обозначается как вторичный проекционно-ассоциативный. Внутренний зернистый слой является местом окончания основной массы проекционных афферентных волокон.
V. Внутренний пирамидный слой, или слой узловых клеток. Состоит в основном из средних и больших пирамидных клеток. Эти нейроны обладают длинными апикальными дендритами, простирающимися вплоть до молекулярного слоя, а также базальными дендритами, распространяющимися более или менее тангенциально по отношению к поверхности. Эти слои четко выражены в передней центральной извилине и незначительно – в других участках коры. Из этого слоя в основном формируются двигательные произвольные пути (проекционные эфферентные волокна).
VI. Слой веретеновидных клеток (полиморфный, или мультиформный). В этом слое расположены преимущественно веретеновидные нейроны, которые имеют короткие извитые верхушечные дендриты, заканчивающиеся в V и IV слоях коры. Аксоны многих клеток слоя объединяются в возвратные волокна, проникая в V слой. Глубинная часть этого слоя переходит в белое вещество (рис. 2.10).
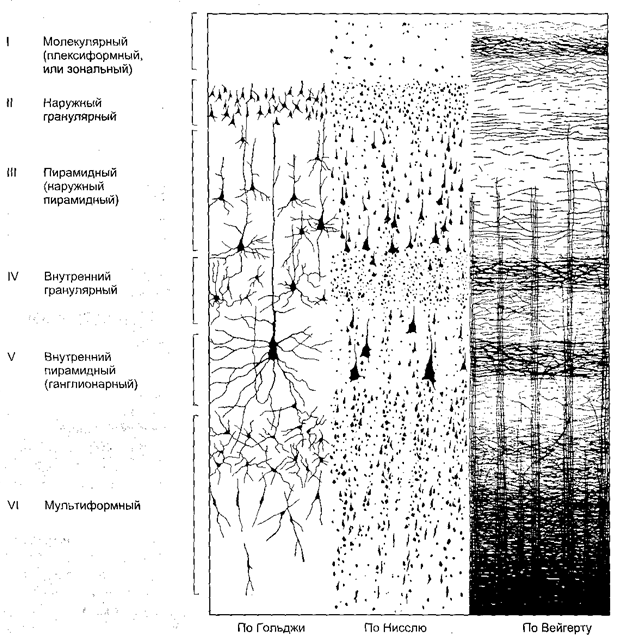
Рис. 2.10. Слоистое строение коры головного мозга
Нужно заметить, что нейроны каждого коркового поля имеют свои особенности строения. Цитоархитектонические слои состоят из нервных и глиальных клеток (олигодендро-, астромакроглии) и многочисленных нервных волокон. Нервные волокна образуют плотные сплетения, называемые нейропилем. Нервные клетки по своей форме весьма разнообразны. Отмечают до 56 разновидностей клеток коры. Более обобщенно выделяют наиболее многочисленные пирамидные нейроны (гигантские Беца, крупные моторные, средние, мелкие), звездчатые и веретенообразные. Доля пирамидных клеток среди всех нейронов коры колеблется от 51 до 86%, звездчатых клеток – от 8 до 47%, веретенообразных нейронов – от 2 до 6% (рис. 2.9).
Функционально в коре выделяют преимущественно возбуждающие нейроны: пирамидные, звездчатые, клетки Мартинотти (перевернутые пирамиды), глиеподобные и преимущественно тормозные: большие корзинчатые, малые корзинчатые, вертикально ориентированные, веретенообразные. Соединения между нейронами обеспечивают многочисленные синапсы и электротонические контакты. Большое значение в деятельности коры имеют шипиковые синапсы. Так, при развитии животных в условиях обогащенной раздражителями среды, по сравнению с сенсорной депривацией, происходит увеличение количества шипиков на дендритах. Умственная отсталость, снижение способности к обучению при хромосомных аберрациях у человека сопровождается уменьшением количества шипиков. Электротонические контакты осуществляются в коре в 20% случаев. Кроме того, в коре описаны несинаптические контакты между нейронами, функциональное назначение такого рода контактов остается еще неясным. В слоях I, II преимущественно имеются дендро-шипиковые контакты, в слоях III, IV – дендро-дендритические и сомато-дендритические, в слое V – сомато-сома-
тические и дендро-дендритические.
Американским физиологом В. Маунткаслом был выдвинут модульный принцип организации нейронов коры. Этот принцип базируется на трех отправных точках.
1. Кора головного мозга состоит из сложных многочисленных ансамблей, основная единица которых образована примерно сотней вертикально связанных нейронов всех слоев коры. Данный ансамбль называется мини-колонкой. В эти мини-колонки входят: а) нейроны, которые получают входные нейроны в основном от подкорковых структур, – например, от специфических сенсорных и моторных ядер таламуса; б) нейроны, получающие входные сигналы от других областей коры; в) все нейроны локальных сетей, образующих вертикальные клеточные колонки; г) клетки, передающие выходные сигналы от колонки назад к таламусу, другим областям коры, а иногда и клеткам лимбической системы.
2. Несколько таких сходных в своей основе простых вертикальных ансамблей могут объединяться с помощью межколоночных связей в более крупную единицу, перерабатывающую информацию, – модуль, или модульную колонку. Несмотря на различную плотность нейронов в слоях разных частей коры, общая структура и функции таких модульных колонок однотипны. Эти колонки различаются лишь по источнику получаемых ими входных сигналов и по мишеням, которым адресуются их выходные сигналы.
3. Маунткасл считает, что модули не только получают и перерабатывают информацию, но и совместно функционируют в составе обширных петель, по которым информация, выходя из колонок, передается другим кортикальным и субкортикальным мишеням, а затем возвращается обратно в кору. Эти петли обеспечивают упорядоченное поступление информации в кортикальные ансамбли.
Связи неокортекса
В новой коре выделяют несколько типов эфферентных и афферентных связей.
Эфферентными волокнами (кортико-фугальными) могут быть:
1) проекционные волокна к подкорковым образованиям (пути: кортико-спинальные, кортико-таламические, кортико-понтийные);
2) ассоциативные волокна, которые идут к одноименным и соседним областям коры одноименного полушария;
3) комиссуральные волокна, которые соединяют области коры обоих полушарий. Основными комиссурами являются мозолистое тело (corpus callosum) и передняя таламическая спайка. Мозолистое тело содержит очень много волокон. Например, у кошек на 1 мм 2 приходится около 700 тысяч волокон.
Афферентными волокнами (кортико-петальными) являются ассоциативные, комиссуральные и таламокортикальный путь – главный афферентный путь к коре от подкорковых образований.
Афферентные волокна заканчиваются в основном в I-IV слоях коры. Исходя из этого, можно предположить, что в процессе обработки информации поверхностные слои ответственны главным образом за восприятие и обработку кортико-петальных сигналов. Особенное значение в этом процессе принадлежит IV слою коры.
Тела важнейших эфферентных нейронов коры лежат преимущественно в более глубоких слоях V-VI. Их считают зоной начала эфферентных путей коры.
(большого мозга), или плащ, cortex cerebri (pallium), - это слой серого вещества полушарий большого мозга , расположенный по его периферии. Морфология коры больших полушарий головного мозга -раздел морфологии нервной системы , предметом исследования которого является макроскопическое и микроскопическое строение коры больших полушарий головного мозга.
Площадь поверхности одного полушария у взрослого человека , как и любая переменная , описывающая живые сущности и явления ,вероятностная переменная величина . Она значительно варьирует по своим значениям . Её средняя величина cоставляет ~2,2·
10 4 мм
2 . При этом на выпуклые (видимые) части извилин приходится одна третья часть всей площади, а на боковые и нижние стенки борозд - две трети.
Наиболее крупные отделы коры головного мозга:
-
древняя кора (палеокортекс),
-
старая кора (архикортекс),
-
новая кора (неокортекс) и
-
межуточная кора.
Основную центральную часть поверхности полушарий составляет новая кора (~95,6% площади поверхности). Старая кора (~2,2% площади поверхности), древняя кора (~0,6% площади поверхности) и межуточная кора (~1,6% площади поверхности) занимают периферию поверхности коры, т. е. расположены по краям плаща большого мозга. Новую кору в связи с ее структурной однородностью называют также изокортексом, а остальные, неоднородные части коры - аллокортексом.
На срезах полушарий большого мозга в некоторых областях (затылочная доля) даже невооруженным глазом видна слоистость коры, характерная для коры в целом . Это чередующиеся серые и белые полосы, ориентированные вдоль поверхности коры. При исследовании с помощью микроскопа видно, что серые полосы составлены по-преимуществу наиболее однородными по своим морфологическим характеристикам нервными клетками , а белые - наиболее однородными нервными волокнами .
Древняя кора у человека и высших млекопитающих состоит из одного слоя клеток, нечетко отделённого от нижележащих подкорковых ядер . Старая кора полностью отделена от подкорковых ядер и представлена чаще всего 2 ÷ 3 слоями клеток. Новая кора может состоять из 6 ÷ 7 слоев клеток. Межуточные формации - переходные структуры между полями старой и новой коры, а также между древней и новой корой могут состоять из 4 ÷ 5 слоев клеток.
Толщина коры, как и любая переменная, описывающая живые сущности и явления,вероятностная переменная. Она варьирует по своим значениям, в различных участках мозга у одного человека и у разных людей толщина коры может изменяться в пределах ~1,5 ÷ 5,0 мм
. Наибольшая толщина наблюдается в верхних участках предцентральной, постцентральной извилин и парацентральной дольки. Обычно толщина коры больше на выпуклой поверхности извилин, чем на боковых поверхностях и на дне борозд.
В состав коры головного мозга входит ~1·
10 9 ÷ 1·
10 10 нейронов и еще большее число глиальных клеток (общее число последних пока не известно). Клетки коры взаимодействуют друг с другом. Одним из средств взаимодействия являются синапсы. Их общее количество в коре оценивается числом ~1·
10 12 . Российский анатом и гистолог В. А. Бец (Бец Владимир Алексеевич, 1834-1894) доказал, что не только вид нервных клеток, но и их взаимное расположение варьирует в различных участках коры. Распределение различных нервных клеток в коре обозначается термином «цитоархитектоника
». Наиболее однородные по своим морфологическим характеристикам нервные клетки располагаются в виде отдельных слоев.
Микроскопические исследования показывают, что в каждом слое, содержатся как тела нервных и глиальных клеток , так и нервные волокна. Эти нервные волокна, составляющие проводящие пути , могут быть отростками нейронов данного слоя коры, нейронов других её клеточных слоев или нейронов других структур мозга. Строение и плотность распределения волокон в различных отделах коры неодинаковы. Распределение нервных волокон в коре головного мозга определяют термином «миелоархитектоника
». Миелоархитектоника коры соответствует её цитоархитектонике.
|
Схема. Слои коры полушарий головного мозга
|
|
|
Примечание : |
Левая вертикальная треть схемы - серебрение препарата по К. Гольджи (Golgi, Camillo, 1843/44-1926, итальянский врач, цитолог, лауреат Нобелевской премии по медицине 1906 г). При такой подготовке препарата окрашивается лишь небольшое число нейронов, но хорошо видны распространение аксонов и апикальных дендритов клеток коры. |
|
Предпосылка
: |
Цитоархитектоника
- молекулярный слой
- наружный зернистый слой
- слой пирамидальных нейронов
- внутренний зернистый слой
- ганглионарный слой
- слой полиморфных клеток
Миелоархитектоника
- полоска молекулярного слоя
- полоска наружного зернистого слоя
- полоска внутреннего зернистого слоя
- полоска ганглионарного слоя.]]
Кора полушарий головного мозга представлена слоем серого вещества толщиной в среднем около 3 мм. Наиболее сильно развита она в передней центральной извилине. Обилие борозд и извилин значительно увеличивает площадь серого вещества головного мозга. В коре содержится около 10-14 млрд нервных клеток. Различные её участки, отличающиеся друг от друга некоторыми особенностями расположения и строения клеток, расположения волокон и функциональным значением, называются полями. Они представляют собой места высшего анализа и синтеза нервных импульсов. Резко очерченные границы между ними отсутствуют. Для коры характерно расположение клеток и волокон слоями.
Типичным для новой коры является наличие шести слоёв, различающихся между собой главным образом по форме входящих в них нервных клеток. При этом на медиальной и нижней поверхностях полушарий сохранились участки старой и древней коры, имеющей 2-слойное и 3-слойное строение. Также выделяется промежуточная кора располагающаяся между старой и новой, а также древней и новой корой. Древняя кора представлена гиппокампом, а старая - участком коры возле обонятельной луковицы на нижней поверхности лобной доли.
Цитоархитектоника
Мультиполярные нейроны коры головного мозга весьма разнообразны по форме. Среди них можно выделить:
- пирамидные
- звёздчатые
- веретенообразные
- паукообразные
- горизонтальные
Пирамидные нейроны составляют основную и наиболее специфическую для коры головного мозга форму. Размеры их варьируют от 10 до 140 мкм. Они имеют вытянутое треугольное тело, вершина которого обращена к поверхности коры. От вершины и боковых поверхностей тела отходят дендриты, заканчивающиеся в различных слоях серого вещества. От основания пирамидных клеток берут начало аксоны, в одних клетках короткие, образующие ветвления в пределах данного участка коры, в других длинные, поступающие в белое вещество.
Пирамидные клетки различных слоёв коры отличаются размерами и имеют разное функциональное значение. Мелкие клетки представляют собой вставочные нейроны, аксоны которых связывают отдельные участки коры одного полушария или двух полушарий. Эти клетки встречаются в разных количествах во всех слоях коры. Особенно богата ими кора головного мозга человека. Аксоны крупных пирамидных нейронов принимают участие в образовании пирамидных путей, проецирующих импульсы в соответствующие центры мозгового ствола и спинного мозга.
Нейроны коры расположены нерезко отграниченными слоями. Каждый слой характеризуется преобладанием какого-либо одного вида клеток. В двигательной зоне коры различают 6 основных слоёв:
- Молекулярный
- Наружный зернистый
- Пирамидальных нейронов
- Внутренний зернистый
- Ганглионарный
- Слой мультиформных клеток
Кора полушарий головного мозга также содержит мощный нейроглиальный аппарат, выполняющий трофическую, защитную, опорную и разграничительную функции.
На медиальной и нижней поверхности полушарий сохранились участки старой, древней коры, которые имеют двухслойное и трехслойное строение.
Молекулярный слой
Молекулярный слой коры содержит небольшое количество мелких ассоциативных клеток веретеновидной формы. Их аксоны проходят параллельно поверхности мозга в составе тангенциального сплетения нервных волокон молекулярного слоя. Основная масса волокон этого сплетения представлена ветвлениями дендритов нейронов нижележащих слоёв.
Наружный зернистый слой
Наружный зернистый слой образован мелкими нейронами диаметром около 10 мкм, имеющими округлую, угловатую и пирамидальную форму, и звёздчатыми нейронами. Дендриты этих клеток поднимаются в молекулярный слой. Аксоны или уходят в белое вещество, или, образуя дуги, также поступают в тангенциальное сплетение волокон молекулярного слоя.
Слой пирамидальных нейронов
Является самым широким по сравнению с другиями слоями коры головного мозга. Он особенно хорошо развит в прецентральной извилине. Величина пирамидных клеток последовательно увеличивается в пределах 10-40 мкм от наружной зоны этого слоя к внутренней. От верхушки пирамидной клетки отходит главный дендрит, который располагается в молекулярном слое. Дендриты, берущие начало от боковых поверхностей пирамиды и её основания, имеют незначительную длину и образуют синапсы со смежными клетками этого слоя. Аксон пирамидной клетки всегда отходит от её основания. В мелких клетках он остаётся в пределах коры; аксон же, принадлежащий крупной пирамиде, обычно формирует миелиновое ассоцативное или комиссуральное волокно, идущее в белое вещество.
Внутренний зернистый слой
В некоторых полях коры развит очень сильно. Однако в других участках он может отсутствовать. Этот слой образован мелкими звёздчатыми нейронами. В его состав входит большое количество горизонтальных волокон.
Ганглионарный слой
Образован крупными пирамидными клетками, причём область прецентральной извилины содержит гигантские клетки, описанные впервые киевским анатомом В. А. Бецем в 1874 году. Они достигают в высоту 120 и в ширину 80 мкм. В отличие от других пирамидных клеток коры гигантские клетки Беца характеризуются наличием крупных глыбок хроматофильного вещества. Их аксоны образуют главную часть кортико-спинальных и кортико-нуклеарных путей и оканчиваются на мотонейронах мозгового ствола и спинного мозга.
Перед выходом из коры от пирамидного пути отходит множество коллатералей. Аксоны от гигантских клеток Беца дают коллатерали, посылающие тормозящие импульсы в саму кору. Также коллатерали волокон пирамидного пути идут в полосатое тело, красное ядро, ретикулярную формацию, ядра моста и нижних олив. Ядра моста и нижних олив передают сигнал в мозжечок. Таким образом, когда пирамидный путь передаёт сигнал, вызывающий целенаправленное движение, в спинной мозг, одновременно сигналы получают базальные ганглии, ствол мозга и мозжечок. Помимо коллатералей пирамидных путей, существуют волокна, которые идут непосредственно от коры к промежуточным ядрам: хвостатому телу, красному ядру, ядрам ретикулярной формации ствола мозга и др.
Слой мультиморфных клеток
Образован нейронами различной, преимущественно веретенообразной формы. Внешняя зона этого слоя содержит более крупные клетки. Нейроны внутренней зоны мельче и лежат на большом расстоянии друг от друга. Аксоны клеток полиморфного слоя уходят в белое вещество в составе эфферентных путей головного мозга. Дендриты достигают молекулярного слоя коры.
Миелоархитектоника
Среди нервных волокон коры полушарий головного мозга можно выделить:
- ассоциативные волокна связывают отдельные участки коры одного полушария
- комиссуральные волокна соединяют кору двух полушарий
- проекционные волокна соединяют кору с ядрами низших отделов центральной нервной системы. Афферентные проекционные волокна заканчиваются в слое пирамидальных нейронов
Кроме тангенциального сплетения молекулярного слоя, на уровне внутреннего зернистого и ганглионарного слоёв расположены два тангенциальных слоя миелиновых нервных волокон и коллатералей аксонов клеток коры. Вступая в синаптические связи с нейронами коры, горизонтальные волокна обеспечивают широкое распространение в ней нервного импульса.
Модуль
I, II, III, IV, V, VI - слои коры
Афферентные волокна
1. кортико-кортикальное волокно
2. таламо-кортикальное волокно
2а. зона распространения специфических таламо-кортикальных волокон
3. пирамидные нейроны
3а. заторможенные пирамидные нейроны
4. тормозные нейроны и их синапсы
4а. клетки с аксональной кисточкой
4б. малые корзинчатые клетки
4в. большие корзинчатые клетки
4г. аксоаксональные нейроны
4д. клетки с двойным букетом дендритов
5. шипиковые звёздчатые клетки, возбуждающие пирамидные нейроны непосредственно и путём стимуляции клеток с двойным букетом дендритов
Исследуя кору больших полушарий головного мозга Сентаготаи и представители его школы установили, что её структурно-функциональной единицей является модуль вертикальная колонка диаметром около 300 мкм. Модуль организован вокруг кортико-кортикального волокна, представляющего собой аксон пирамидной клетки III слоя того же полушария, либо от пирамидальных клеток противоположного. В модуль входят два таламо-кортикальных волокна специфических афферентных волокна, оканчивающихся в IV слое коры на шипиковых звёздчатых нейронах и отходящих от основания дендритах пирамидальных нейронов. Каждый модуль, по мнению Сентаготаи разделяется на два микромодуля диаметром менее 100 мкм. Всего в неокортексе человека примерно 3 млн модулей. Аксоны пирамидальных нейронов модуля проецируются на три модуля той же стороны и через мозолистое тело посредством комиссуральных волокон на два модуля противоположного полушария. В отличие от специфических афферентных волокон, оканчивающихся в IV слое коры, кортико-кортикальные волокна образуют окончания во всех слоях коры, и, достигая I слоя, дают горизонтальные ветви, выходящие далеко за пределы модуля.
Помимо специфических афферентных волокон, на выходные пирамидальные нейроны возбуждающее влияние оказывают шипиковые звёздчатые нейроны. Различают два типа шипиковых клеток:
- шипиковые звёздчатые нейроны фокального типа, образующие множественные синапсы на отходящих от верхушки дендритах пирамидального нейрона
- шипиковые звёздчатые нейроны диффузного типа, аксоны которых широко ветвятся в IV слое и возбуждают базальные дендриты пирамидальных нейронов. Коллатерали аксонов пирамидных нейронов вызывают диффузное возбуждение соседних пирамид.
Тормозная система модуля представлена следующими типами нейронов:
- клетки с аксональной кисточкой образуют в I слое множественные тормозные синапсы на горизонтальных ветвях кортико-кортикальных волокон
- корзинчатые нейроны - тормозные нейроны, образующие тормозящие синапсы на телах практически всех пирамидных клеток. Они подразделяются на малые корзинчатые нейроны, оказывающие тормозящее влияние на пирамидные нейроны II, III и V слоёв модуля, и большие корзинчатые клетки, располагающиеся на периферии модуля и имеющие тенденцию подавлять пирамидные нейроны соседних модулей
- аксоаксональные нейроны, тормозящие пирамидные нейроны II и III слоёв. Каждая такая клетка образует синапсы на начальных участках аксонов сотен нейронов II и III слоёв. Они тормозят, таким образом, кортико-кортикальные волокна, но не проекционные волокна нейронов V слоя.
Система угнетения тормозных нейронов:
- клетки с двойным букетом дендритов располагаются во II и III слоях и, угнетая тормозные нейроны, производят вторичное возбуждающее действие на пирамидные нейроны. Ветви их аксонов направлены вверх и вниз и распространяются в узкой колонке. Таким образом, клетка с двойным букетом дендритов растормаживает пирамидные нейроны в микромодуле.
Мощный возбуждающий эффект фокальных шипиковых звёздчатых клеток объясняется тем, что они одновременно возбуждают пирамидные нейроны и клетку с двойным букетом дендритов. Таким образом, первые три тормозных нейрона тормозят пирамидные клетки, а клетки с двойным букетом дендритов возбуждают их, угнетая тормозные нейроны.
Однако, также существуют критические и альтернативные концепции, ставящие под сомнение модульную организацию коры больших полушарий и мозжечка. Безусловно, влияние на эти воззрения оказало предсказание в 1985 г. и в дальнейшем открытие в 1992 г. диффузного объёмного нейротрансмиттинга.
Резюме
Межнейрональные взаимосвязи нейронов коры больших полушарий головного мозга можно представить следующим образом: входящая информация поступает из таламуса по таламо-кортикальным волокнам, которые заканчиваются на клетках IV слоя. Его звёздчатые нейроны оказывают возбуждающее воздействие на пирамидные клетки III и V слоёв, а также на клетки с двойным букетом дендритов, которые блокируют тормозные нейроны. Клетки III слоя образуют волокна, которые связывают между собой различные отделы коры. Клетки V и VI слоёв формируют проекционные волокна, которые уходят в белое вещество и несут информацию другим отделам центральной нервной системы. Во всех слоях коры находятся тормозные нейроны, играющие роль фильтра путём блокирования пирамидных нейронов.
Кора различных отделов характеризуется преимущественным развитием тех или иных её слоёв. Так, в двигательных центрах коры, например в передней центральной извилине, сильно развиты III, V и VI и плохо выражены II и IV слои. Это так называемый агранулярный тип коры. Из этих областей берут начало нисходящие проводящие пути центральной нервной системы. В чувствительных корковых центрах, где заканчиваются афферентные проводники, идущие от органов обоняния, слуха и зрения, слабо развиты слои, содержащие крупные и средние пирамидные клетки, тогда как зернистые слои достигают своего максимального развития. Это гранулярный тип коры.
| Мозжечок |