Позвоночные животные.
Высший ствол «древа животных » образуют позвоночные животные. От одного из разветвлений этого ствола произошел человек. Только у позвоночных животных имеется нервная система, сходная в основных своих чертах с нервной системой человека.
Вследствие этого анатомического сходства могут явиться также и сходства психические. У низших позвоночных животных психическое их сходство с человеком незначительно, но у млекопитающих оно все вырастает по мере того, как мозг млекопитающего приближается к мозгу человека.
Центральная нервная система позвоночных животных состоит из головного и спинного мозга, В восходящем ряду позвоночных головной мозг обнаруживает все более развитое строение, и каждый класс позвоночных имеет головной мозг своего особого устройства и формы. Так, например, мозг земноводных, или амфибий, развит не так высоко, как мозг пресмыкающихся, или рептилий; а мозг птиц оказывается еще более высоко развитым, нежели мозг рептилий. Мозг животных млекопитающих вообще стоит выше мозга рептилий, но и в пределах группы млекопитающих, в свою очередь, наблюдается весьма значительный рост степени совершенства мозга при сравнении низших представителей группы с высшими.
Ступенчатая последовательность все более развитых форм мозга млекопитающих отмечает таким образом ряд и психических ступеней, почему мы и должны заняться сначала рассмотрением устройства этих анатомических форм. Мозг позвоночных животных состоит из следующих частей: 1) двух полушарий большого головного мозга, 2) промежуточного мозга, 3) среднего мозга, 4) мозжечка, 5) продолговатого, или заднего мозга.
Рисунок 6.1. Мозг лягушки и мозг рыбы
Рис. 7. Мозг лягушки
Рис. 8. Мозг рыбы (форели)
От полушарий большого мозга берут начало обонятельные нервы. Большой мозг у амфибий еще не особенно велик (Рис. 6.1 , рис. 7, g), у рептилий он уже значительно больше (Рис. 6.2), еще больше он у птиц (Рис. 6.2) и наивысшего своего развития он достигает у млекопитающих (Рис. 6.3).
От промежуточного мозга берут начало зрительные нервы. К промежуточному мозгу относится зрительный бугор (Thalamus opticus). У амфибий промежуточный мозг лежит позади большого мозга (Рис. 6.1). Но у рептилий и у птиц большой мозг распространяется назад настолько далеко над промежуточным, что совсем закрывает собой промежуточный мозг, так что последнего сверху уже не видно (Рис. 6.2). Остается заметным лишь маленький вырост промежуточного мозга, называемый эпифизой, или верхней мозговой железой.
Средний мозг у большинства позвоночных образует две выпуклости (Рис. 6.1). У рептилий эти выпуклости достигают значительных размеров (Рис. 6.2). У птиц они разделяются мозжечком (Рис. 6.2). У млекопитающих средний мозг разделен на четыре части, называемые четверохолмием (Рис. 6.3 , рис. 11, m); у высших млекопитающих часть эта мала и незначительна, четверохолмие здесь закрыто сверху большим мозгом (Рис. 6.3).
Мозжечок у амфибий невелик (Рис. 6.1); у рептилий он несколько больше (Рис. 6.2); у птиц (Рис. 6.2) и млекопитающих (Рис. 6.3) он уже является высоко развитым, так как полет птиц и бег млекопитающих нуждаются в сложной регуляции со стороны нервных центров, локализующихся в мозжечке.
Задний мозг образует продолжение спинного мозга, почему и обозначается как продолговатый мозг (Рис. 6.1 , рис. 7, n). От него отходит в разные стороны множество
Здесь не представляется возможным рассмотреть все ступени постепенного развития мозга в ряду позвоночных.
Предметом нашего рассмотрения будут только четыре класса: рыбы, амфибии, птицы, млекопитающие.
Рыбы.
Из рыб мы рассмотрим лишь тех, которые относятся к классу костистых (Teleostei); сюда войдут наиболее известные рыбы (карп, форель, щука, окунь и т. д.). Мозг этих рыб обладает некоторыми отличиями, которых не имеется в такой же форме у низших рыб (напр., у акул). Первый отдел мозга млекопитающих - большой мозг - здесь имеет сравнительно небольшие размеры; его верхняя стенка (Pallium) весьма тонка, тогда как средний мозг поразительно велик (Рис. 6.1). Жизнь рыб регулируется главным образом инстинктами, но наряду с ними у многих рыб имеется также и память; отсюда можно видеть, что образование путей, складывающихся в процессе индивидуальной жизни, может происходить не только в верхней стенке большого мозга, но также и в других частях мозга. Существование памяти у рыб доказано многочисленными опытами . Многие рыбы, живущие в аквариумах, могут становиться ручными и подплывают к человеку, приносящему им корм. Один господин в Майнце так приучил радужную форель, что она из его рук брала корм, но после того, как этот человек однажды на секунду вытащил рыбку из воды за хвост наружу, рыбка в течение трех дней избегала приближаться к корму. Установлена способность многих рыб ориентироваться в пространстве; так, напр., колюшки снова отыскивают свое гнездо на пространстве радиусом в 10 метров; некоторые рыбы (щуки, форели) возвращаются к «стоянкам » , где они подстерегают добычу, с довольно значительных расстояний (до 6 километров) . Неоднократно замечалось, что рыбы в пруду запоминают внешний вид человека, регулярно носящего им корм; так, напр., в одном питомнике для форелей сторож, приносивший корм, обыкновенно являлся в одеянии ярко-красного цвета, и вот всякому, кто надевал эту одежду, тоже удавалось подманивать к себе форелей.
М. Окснер произвел с морскими рыбами (Coris julis, Serranus scriba) ряд опытов, с несомненностью доказывающих существование у рыб памяти. Он вешал в аквариуме разноцветные цилиндры и замечал, что рыбы искали пищи в том цилиндре, в котором он их кормил раньше .
Подобным образом К. ф.-Фриш установил, что рыбу-скомороха (Phoxinus laevis) можно приучить брать корм из кормушки определенного цвета; при этом оказалось, - по вопросу о различении цветов, что хотя эти рыбы неотчетливо различают красный и желтый цвет, однако зеленый от синего, равно как и оба эти последние цвета от красного и желтого, они отличают вполне хорошо .
Амфибии.
Переходя от рыб к земноводным, мы видим, что у них передний мозг, промежуточный мозг, средний и задний мозг хорошо развиты, тогда как мозжечок развит незначительно (Рис. 6.1). Инстинкты земноводных лишь в самой незначительной доле локализованы в переднем мозгу; нервные центры, с которыми они связаны, находятся главным образом в следующих частях головного мозга и в спинном мозгу. Если вырезать у лягушки передний мозг, то большая часть ее жизненных отправлений сохраняется, по сообщению физиолога Шрадера . Оперированные таким образом животные самостоятельно едят, с наступлением зимы погружаются в ил на дне водоема, с приходом весны выходят на поверхность и спариваются. «Я не в состоянии, - писал названный ученый, - путем простого наблюдения отличить - среди нормальных и оперированных лягушек, живущих в лягушином пруду Физиологического института - лягушек, лишенных переднего мозга и совершенно оправившихся после операции; последних выдает только тонкая линия рубца на коже головы или ощутительный дефект черепной крышки. Лягушка, лишенная переднего мозга, плавает, прыгает, бегает с той же ловкостью, как и нормальная особь, и она поразительно хорошо ориентируется в пространстве с помощью чувства зрения » .
Сравнительно с инстинктами, память играет у земноводных лишь мало заметную роль: память не отсутствует у них вполне, - как в том легко может убедиться всякий, содержащий в неволе древесную лягушку, - но эта память так незначительна, что я здесь на ней останавливаться не буду.
Птицы.
Но у птиц память играет большую роль; память локализована у птиц главным образом в переднем мозгу. Если у голубя или у другой птицы вырезать передний мозг (не поранив при этом промежуточного мозга со зрительными нервами), то животное становится «душевно-слепым » , т.-е. оно теряет понимание предметов внешнего мира.
Рисунок 6.2. Мозг рептилии и мозг птицы

Рис. 9. Мозг рептилии (крокодила)
Рис. 10. Мозг птицы (голубя)
Оперированная птица, правда, в совершенстве сохраняет способность бегать и без труда летит к месту обычного отдыха, но предметы, которые эта птица видит, она уже больше не узнает. «Она не делает никакого различия - отражается ли на ретине ее глаза какой-либо неодушевленный предмет или же собака, кошка, хищная птица, или же сачок, которым она была поймана; для оперированной птицы все эти предметы лишь препятствия, которые она стремится обойти, через которые хочет перелезть, или которыми желает воспользоваться, как местом для отдыха при перелетах » .
Голубь без переднего мозга дает себя поймать, не обращаясь в бегство; сокол без переднего мозга дает себя схватить, не обороняясь. Голубь без переднего мозга в брачный период следует своему инстинкту, усердно воркует и проявляет все движения ухаживающего самца, но самка не воспринимается им как таковая, и он остается по отношению к ней совершенно безразличным.
«Как оперированный самец не обнаруживает более никакого интереса к самке, так и оперированная самка не интересуется своими детенышами; едва оперившиеся птенцы преследуют мать непрестанными криками, и эффект получается тот же, как если бы они обращались к камню » .
Голуби и куры без переднего мозга перестают замечать свою пищу и могут умереть с голоду на куче зерна; чтобы поддержать их жизнь, необходимо вкладывать им зерна в клюв. Здесь уместно вспомнить факт, упоминавшийся выше, а именно: куры путем индивидуального опыта научаются распознавать годную для них пищу. Таким образом, в процессе еды оживают вспоминаемые впечатления, связанные с нервными путями, возникшими в индивидуальной жизни и проходящими через передний мозг. С удалением переднего мозга выпадают и эти образы воспоминаний.
На наблюдениях Макса Шрадера особенно отчетливо можно видеть, как в процессе еды хищных птиц вступают во взаимодействие инстинктивные побуждения и индивидуально приобретенный опыт. Шрадер брал из гнезда молодых неоперившихся соколят. Когда соколятам показывали двигающуюся мышь, они тотчас протягивали лапы к добыче, громко крича, и крепко цеплялись за нее когтями, хотя птенцы еще совершенно не были в силах причинить мыши какой бы то ни было вред; они даже ни разу не сделали попытки склюнуть добычу, и их приходилось еще кормить из рук; кусок лошадиного мяса, обрывок платка, палец человека встречали такой же прием. Таким образом, поводы к схватыванию дает, повидимому, движущееся тело. В нормальных условиях соколята вскармливаются родителями, которые дают им живую добычу и постепенно учат птенцов ее распознавать. Но в данном случае у подопытных птенцов это не имело места; им давали в пищу только лошадиное мясо и позже - мертвых голубей. «Когда они совершенно оперились, в их клетку опять впустили живую добычу - белых мышей; интересно было наблюдать, как соколята сначала издали наблюдали пришельцев-мышей, как затем стали осторожно придвигаться и ходить вокруг спокойно грызущих мышей, осматривая их со всех сторон; некоторые птенцы затем опять отошли и сели на нашесте; один птенец боязливо протянул лапу к мыши, но едва коснулся ее, как быстро отдернул лапу назад; мышь отскочила в сторону, сокол убежал далеко прочь, в большем страхе, чем мышь. Опыт был повторен, соколы становились все смелее, но только через два-три дня мыши были пойманы и с"едены соколами » . Из этого описания видно, что хотя у соколят и было врожденное влечение к схватыванию живой добычи, однако они лишь в процессе опыта узнали, что мыши являются такой добычей, которую можно схватывать и есть .
Млекопитающие.
Так как об инстинктах и об уме птиц мы уж достаточно говорили в других местах этой книги, то мы перейдем теперь к млекопитающим животным.
Три части мозга являются у млекопитающих в особенно развитом виде: передний мозг, мозжечок и задний мозг. Промежуточный мозг, напротив, является погруженным под передним мозгом, и его можно видеть снаружи лишь с нижней стороны мозга. Средний мозг, образующий четверохолмие, теряет свое значение; у низших млекопитающих его еще можно видеть позади переднего мозга (рис. 11). У высших млекопитающих средний мозг является настолько малым, что совершенно закрывается сверху разросшимся передним мозгом (рис. 12).
Переднему мозгу мы должны уделить особое внимание, так как он является главным седалищем памяти и рассудка. Наибольшая часть нервных связей, образующихся в индивидуальной жизни, проходит через передний мозг. При удалении переднего мозга подвергаются выпадению все те проявления, на основании которых можно заключать о присутствии памяти, интеллекта и размышления.
Рисунок 6.3. Мозг кролика и мозг собаки
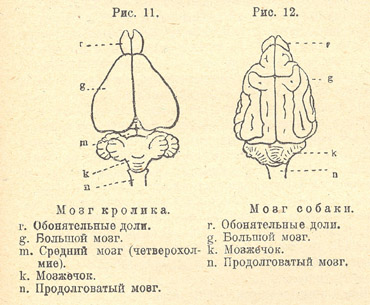
Рис. 11. Мозг кролика
Рис. 12. Мозг собаки
Собака с вырезанным передним мозгом так же, как и голубь с вырезанным передним мозгом, оказывается «душевно-слепой » , т.-е. она не замечает ни кормушки с водой ни угрозы хлыстом, она не узнает даже поставленной перед ней кошки. Эта собака не реагирует ни на ласковые ни на грубые слова и остается равнодушной, даже когда ее гладят, так как у нее исчезли психические связи с окружающими людьми . Но такая собака может еще долгое время оставаться в живых, так как она в состоянии есть и пить, и большая часть ее рефлексов сохранена.
У низших млекопитающих передний мозг является сравнительно небольшим и еще не имеет извилин. Так, у насекомоядных, у летучих мышей и у большинства сумчатых поверхность мозга представляется гладкой, то же имеется и у большинства грызунов. Но у высших млекопитающих кора мозга увеличивается в своих размерах, и на ней появляются углубляющиеся извилины. Извилистую мозговую кору имеют все хищные животные и все копытные, равно как и тюлени, дельфины и киты. Из обезьян низшие породы имеют немного извилин, высшие обезьяны обладают более сложной системой извилин.
По внешнему виду извилин мозга, т.-е. по роду ветвления и по направлению извилин мозговой коры, антропоидные (человекообразные) обезьяны ближе всего стоят к человеку. У рода гиббонов (Нуlobates) можно видеть только главные борозды, но у шимпанзе, оранг-утанга и гориллы имеется уже сложная система извилин, весьма сходная с таковой человека. Что же касается массы мозга и, следовательно, его веса, то здесь наблюдаются большие различия. Мозг шимпанзе весит 350 - 400 граммов, мозг гориллы - около 425 гр., тогда как вес мозга человека у низших рас равняется 900 - 1.000 гр., а у высших достигает 1.300 - 1.500 гр. и более.
При этом следует еще обратить внимание на толщину мозговой коры и на число нервных клеток. Существуют такие млекопитающие, у которых нервные клетки расположены слоями, далеко отстоящими один от другого. Эти животные имеют 5.000 - 10.000 клеток на один куб. мм. Таким мозгом обладают сумчатые, неполнозубые, жвачные, слоны и киты. Затем более тесное расположение клеток мы находим у хищников и у тюленей: от 15.000 - 20.000 клеток на один куб. мм. Наиболее тесным расположением клеток отличаются грызуны, полуобезьяны и обезьяны: 35.000 - 50.000 на один куб. мм. Человек имеет весьма тесное расположение нервных клеток. Высокая степень разумности, присущая человеку, об"ясняется, таким образом, величиной переднего мозга, сложностью системы извилин, - что дает увеличение площади коры, - и тесным расположением клеток в мозговой коре - следовательно, наличностью большего количества нейронов.
─────── |
В зоологии класс млекопитающих делится на три подкласса: клоачные (monotremata), (утконос и ехидна), сумчатые (marsupialia), плацентарные (placentalia), при чем сумчатые животные разделяются на несколько отрядов, а плацентарные на еще большее число отрядов (наприм., неполнозубые, насекомоядные, грызуны, копытные, китообразные, хищные, тюлени, летучие мыши, полуобезьяны и обезьяны). Но, с точки зрения зоопсихологической, целесообразно соединить в одну группу всех тех млекопитающих, мозг которых в своем устройстве сохраняет черты примитивного типа; и, с другой стороны, следует отдельно поставить группу высших млекопитающих, обладающих мозгом более сложного строения .
Примитивный тип мозга мы встречаем у клоачных и сумчатых, а среди плацентарных животных - у грызунов, насекомоядных и неполнозубых. Передний мозг у них еще не достиг значительного развития, он чаще всего еще не покрывает собою среднего мозга (четверохолмие) (рис. 11). На нижней стороне переднего мозга имеются широкие обонятельные лопасти, от которых отходят сильно развитые обонятельные нервы; значительная часть переднего мозга оказывается, таким образом, занятой обонятельными долями. Поверхность коры переднего мозга гладка, извилин не имеет, и мозговая кора еще не достигла значительного развития . В умственном отношении эти животные стоят невысоко, но инстинкты их достигли большого совершенства, ибо эти животные оказываются способными производить удивительные сооружения (напомним, например, постройки бобров, гнезда орешниковой сони и белки, подземные норы крота, хомяка и т. д.).
Слабое развитие умственных способностей этих млекопитающих сказывается еще в том, что дрессировкой их удается достигнуть весьма немногого; никогда не показывают дрессированных ежей, крыс или кроликов . Но все же у этих животных имеется память на места и также общая способность запоминания. Поэтому они легко поддаются приручению. В Триесте я видел в одном ресторане ежа, который вечером постоянно бегал среди посетителей и позволял себя кормить. У меня самого в квартире жил молодой еж, которого вечером выпускали из его ящика, и который тогда начинал оживленно бегать по комнате и ел из рук. Для своих опытов по наследственной передаче признаков я уже много лет держу почти ручную расу крыс; когда крысы вырастают в ящике так, что их никто не касается, то они боятся человека и не любят, чтобы их брали руками; если же крыс с самой ранней поры брать на руки, то они приручаются настолько, что я мог позволять с ними играть моим детям.
Мы переходим теперь к тем млекопитающим, мозг которых представляется изборожденным извилинами, и мозговая кора которых оказывается хорошо развитой. Таким образом, мы подходим к высшим ступеням умственной деятельности животных. Извилистой мозговой корой обладают хищники (рис. 12), тюлени, копытные, китовые (дельфины и киты), а также обезьяны. В каждом из этих отрядов животных существует особенный тип расположения извилин, чем свидетельствуется, что извилины возникли в каждом отряде независимо, т.-е. что в порядке филогенетического развития в каждом из этих отрядов первоначально имелись формы с мозгом без извилин. В филогенетическом ряду приматов можно еще и теперь видеть как бы ряд различных ступеней пройденного развития: полуобезьяны имеют, по большей части, мозг, лишенный извилин; в мозгу низших обезьян (например, у игрунковой обезьяны - Hapale leoninus) только лишь намечаются первые следы извилин; от этих форм вверх идет все нарастающее усложнение борозд вплоть до прекрасно развитой системы извилин мозга человекообразных обезьян; мозг этих последних имеет уже очень близкое сходство с мозгом человеческим.
Все те млекопитающие животные, которые уже исстари пользуются репутацией умных, как, напр., слон, лошадь, собака, лисица и большая часть обезьян, - равно как все те животные, у которых с помощью дрессировки удается достигнуть значительных результатов (собаки, львы, лошади, слоны, морские львы и обезьяны), - все они имеют весьма извилистый передний мозг. О проявлении ума антропоморфных обезьян мы уже упоминали. При посредстве нового метода стуков еще точнее было узнано, как высоко развиты психические способности у лошадей и собак. Об этом уже шла речь выше, и поэтому мне нет нужды останавливаться здесь снова на уме млекопитающих.
Следует еще только упомянуть, что применительно к инстинктивным чувствованиям млекопитающих позволительно пользоваться теми же терминами, какие мы применяем к человеку. Так, напр., когда речь идет о собаке, мы можем говорить о радости или печали, о симпатии или антипатии, о тоске и ревности, об испуге и страхе, о гневе и ненависти. Так и говорят обычно в обыденной речи, а отмеченные нами аналогии в строении мозга человека и высших животных придают еще и научное обоснование означенному словоупотреблению.
»Преувеличение: в цирках мыши, крысы, кролики являются многократно выполнителями самых причудливых «номеров » . (Прим. ред.).
§ 37. Ассоциативный центр мозга рептилий Рассмотрев общий план строения нервной системы, следует отдельно остановиться на новых принципах организации и работы мозга, впервые реализованных у рептилий. Нервная система архаичных амниот стала логическим развитием строения удачной амфибийной конструкции. Однако мозг амфибий практически выполнял функцию сложного рефлекторного аппарата, а его интеллектуальные возможности остались невостребованными. Эволюция амфибий решалась мышцами, зубами, линейными размерами и масштабами
Рис. III-8. Сагиттальный (а) и горизонтальный (б) гистологические срезы через голову ящерицы (Lacerta agilis ) в конце эмбрионального развития.
Относительный размер мозга в несколько раз больше, чем у амфибий. К моменту вылупления рептилии обладают эффективным вестибулярным аппаратом, зрением, слухом и обонятельной системой.Размножения. Шло элементарное освоение пищевых ресурсов, где для развития сложного поведения не было ни места, ни биологической необходимости. Со следами этого периода эволюции позвоночных мы сталкиваемся, пытаясь выработать условные рефлексы у различных представителей современных амфибий. Крайне низкая обучаемость и отсутствие долговременной памяти для накопления индивидуального опыта показывают, что сложных поведенческих задач перед древними амфибиями никогда в их эволюции не стояло. Особенности развития органов чувств и признаки сложного поведения рептилий базируются на особенностях структурной организации головного мозга. Мозг рептилий отличается от мозга амфибий как в количественном, так и в качественном отношении. До появления амниот стратегии поведения или реакция на конкретный раздражитель выбирались по принципу доминантности (см. рис. III-6, е ). Этот принцип состоит в том, что выраженного крупного ассоциативного центра мозга у многих первичноводных позвоночных или амфибий нет (см. рис. III-6, e). Выбор формы поведения происходит на основании сравнения активностей примерно равноценных отделов мозга, обслуживающих различные органы чувств. Решающее значение играет уровень возбуждения мозговых аналитических центров одного из анализаторов. Представительство органа чувств, достигшее в мозге наибольшего возбуждения, и становится основной областью для принятия решения. После выбора одной из инстинктивных реакций происходит её поведенческая реализация. Этот процесс осуществляется под контролем того же простого сравнения доминантностей. Если в процессе осуществления реакции возникает новое раздражение, которое изменяет соотношение возбуждений органов чувств, то поведенческая реализация инстинктивного процесса останавливается. Каждая конкретная ситуация отличается от предыдущей, но задействуется тот же набор органов чувств. Если наибольшее возбуждение достигается в той же сенсорной системе, то поведение сохраняется, а если в другой, то изменяется. Поскольку абсолютно идентичные условия в естественной жизни практически не встречаются, поведение даже самых примитивных анамний будет бесконечно разнообразно. Следовательно, поведение каждой особи будет индивидуальным с довольно высокой динамической адаптивностью. Первые признаки ассоциативного центра появились ещё в мозге амфибий. У них таким центром мог стать средний или промежуточный мозг. Для этого были все основания. В промежуточном мозге находятся нейроэндокринные центры, контролирующие половое поведение, миграции и энергетический баланс организма анамний. Через
Возникновение мозга птиц
Птицы представляют собой обособленный класс теплокровных, двуногих и обычно летающих амниот. Они покрыты специализированными роговыми придатками - перьями. Современные птицы лишены зубов, но обладают удлинёнными челюстями, которые образуют клюв. Наземное передвижение обеспечивают парные задние конечности. Передние конечности видоизменены в крылья, приспособленные для полёта. Большинство птиц способны летать. Они могут находиться в воздухе очень долго и перелетать на большие расстояния. Сезонные миграции могут составлять несколько тысяч километров. В стабильном климате многие птицы не мигрируют или перемещаются на небольшие расстояния. Летают не все птицы. Пингвины, киви и страусы не обладали этой способностью или утратили её вторично. Птицы - яйцекладущие животные. Все птицы откладывают яйца, но не все их насиживают и заботятся о птенцах. Для нормального развития зародышей необходима контролируемая температура окружающей среды. Это достигается насиживанием яиц, устройством защищённых гнёзд или созданием специальных конструкций, где температура поддерживается за счёт химических реакций распада. Поведение птиц крайне сложно и разнообразно, как и их внешний вид (рис. III-11). У птиц в разных пропорциях встречается инстинктивное и ассоциативное поведение и сохраняется способность к обучению на протяжении всей жизни. Многие птицы обладают хорошей памятью и могут осуществлять несложную инструментальную деятельность. Они легко обучаются решать простые задачи и подражать человеческому голосу. «Слова», произносимые птицами, являются эмоциональным символом ситуационного настроения, в котором они их запомнили, а не результатом рассудочной деятельности. В отличие от всех рассмотренных выше групп животных, птицы любят играть друг с другом или с различными предметами. Они первые среди животных смогли заниматься деятельностью, не приносящей непосредственной биологической выгоды. Мозг птиц обеспечивает возможность предвидения событий. Они рассчитывают своё поведение на несколько шагов вперёд, что уже характерно для высших млекопитающих и человека. Это сходство сложного поведения с поведением млекопитающих свойственно относительно небольшой группе попугаев, врановых, хищных и некоторых других видов. Поведение большинства птиц контролируется преимущественно врождёнными
Рис. I II- 11. Разнообразие птиц.
Столь подробное описание морфологических особенностей строения продиктовано уже упомянутой парадоксальной специализацией птиц. Скелет, покровы, кровеносная, выделительная и дыхательная системы имеют столько специфических особенностей, что их вполне хватило бы для гарантированного исчезновения любой другой группы позвоночных. Исключение составляет нервная система птиц. Во всех систематических группах птиц она построена удивительно однотипно. Анатомически различия обычно сводятся к размерам головного мозга (рис. III-12), а видовые особенности выявляются только на цитоархитектоническом уровне. Она столь же специализирована, как и другие органы, но обладает уникальным архетипом строения, который оказался пригодным для всех форм адаптивного поведения и сред обитания. § 43. Нервная система и органы чувств птиц Нервная система птиц состоит из центрального и периферического отделов. Головной мозг птиц крупнее, чем у любых современных представителей рептилий. Он заполняет полость черепа и имеет округлую форму при небольшой длине (см. рис. III-12). Самый крупный отдел - передний мозг. Он состоит из двух полушарий с гладкой поверхностью или слабо обозначенными продольными углублениями. Эти углубления не являются истинными бороздами, а отражают границы слоёв внутренних ядер (рис. III-13, а , в ). Только у попугаев можно заметить небольшое морфологическое обособление височной доли полушарий. Основной объём полушарий занимают подкорковые ядра, кора имеет рудиментарное строение и занимает небольшую часть верхней стенки мозга. Полушария переднего мозга простираются назад до контакта с мозжечком. Следствием разрастания переднего мозга назад, а мозжечка - вперёд является то, что промежуточного мозга снаружи совсем не видно, хотя его можно определить по выросту эпифиза. Эпифиз у птиц развит незначительно, а гипофиз достаточно крупный. Средний мозг сильно развит, но из-за бокового расположения имеет нехарактерную внешнюю морфологию. Передние выпячивания крыши среднего мозга сдвинуты латерально. Их часто называют зрительными долями (Lobi optici). Мозжечок состоит из массивной средней части (червя), пересекаемой обычно 9 извилинами, и двух небольших боковых долей, которые гомологичны клочку мозжечка млекопитающих. Задний и продолговатый мозг имеет два резких изгиба, обусловленных ориентацией и подвижностью головы птиц. Гистологическое строение головного мозга птиц существенно отличается от такового у других групп позвоночных. Уже у амфибий можно выявить старый и древний стриатум, септум и в зачаточном виде базальные ядра переднего мозга. У рептилий появляется новый
Рис. III-12. Головной и спинной мозг птиц.
Головной мозг птиц обладает крайне стабильной анатомией. Он укорочен в рострокаудальном направлении, а на его поверхности видны только парные полушария переднего мозга, крупный мозжечок и передняя часть крыши среднего мозга. Внешняя форма головного мозга практически не отличается даже у отдалённых видов. а, в - хохлатый осоед (Pernis ptilorhynchus ); б - серый гусь (Anser anser ); г - павлин (Pavo cristatus ).Стриатум, который становится доминирующей структурой переднего мозга (см. рис. III-12; III-13, а). Важно подчеркнуть, что новый стриатум возникает у птиц вопреки развитию зачаточного неокортекса рептилий. Иначе говоря, у рептилий получили значительное развитие структуры стриатума и септума переднего мозга. Они расположены в базальных частях переднего мозга птиц и развиты намного лучше, чем у рептилий (см. рис. III-13,а ). Однако рептилии приобрели и зачаточные корковые структуры переднего мозга, которые сформировались в результате развития дополнительной (половой) обонятельной системы. Эти структуры стали выполнять у рептилий функции нового интегративного мозгового центра на базе переднего мозга. Казалось бы, дальнейшее развитие корковой системы переднего мозга гарантировало бы птицам необходимые поведенческие преимущества. Тем не менее этого не произошло. У птиц корковые структуры, впервые появившиеся в переднем мозге рептилий, носят откровенно рудиментарный характер. Эволюция корковых структур переднего мозга рептилий полностью остановилась у птиц. Зачатки палео-, архи- и неокортекса практически не играют роли в контроле поведения, поскольку обоняние у птиц развито намного меньше, чем у рептилий. У большинства современных птиц нет развитого обоняния, а вомероназальная система полового обоняния у них вообще отсутствует. По-видимому, в период отделения предков птиц из общего рептилийного ствола обоняние перестало играть для них какую-либо роль. Крупный передний мозг остался функционально невостребован и стал морфологическим субстратом для возникновения ассоциативных центров. Зачаточные корковые структуры не могли играть существенной роли на фоне «освободившихся» от своих функций огромных ядер стриатума и септума. Невостребованная нейронная система этих структур надолго обеспечила мозг птиц резервами памяти и возможностями развития сложного адаптивного поведения. Следы кортикальных структур переднего мозга птиц расположены только в дорсальной зоне полушария, а большую часть мозга занимают стриатум, септум и неостриатум. Функции ассоциативного центра в переднем мозге птиц выполняют чрезвычайно развитые структуры стриатума (рис. III-14, а ). Однако историческое название «гиперстриатум» не отражает реального происхождения этого центра птиц. Ранее считалось, что гиперстриатум возник из стриатума и является его гомологом. Специальные исследования кинетики пролиферации и миграции нейробластов у птиц позволили установить, что гиперстриатум состоит из нейронов различного происхождения.
Рис. III-13. Цитоархитектоника головного мозга птиц на примере обыкновенной неясыти (Strix aluco ).
Буквы сечений на схеме мозга соответствуют обозначениям гистологических срезов. Микрофотографии гистологических срезов сделаны в соответствии с линиями, показанными на цветной схеме мозга с латеральной поверхности. Оранжевый цвет - передний мозг, красный - средний мозг, фиолетовый - мозжечок, синий - задний и продолговатый мозг.
Они по большей части мигрируют из латеральной (paleopallium ) и новой коры (neopallium ). Особенностью развития стриатума стало формирование структур ядерного типа, а не стратифицированных корковых образований. У птиц и рептилий разрастание стриатума приводит к экспансии дорсального внутрижелудочкового бугорка, который практически полностью занимает полость латеральных желудочков (см. рис. III-14, а ). Гиперстриатум птиц (рептилии имеют его признаки) выполняет функции новой коры. Он представляет собой высший ассоциативный центр, который определяет принятие решений и является основной зоной хранения индивидуального опыта. Через стриатум осуществляются контроль за двигательными функциями и связь с лимбической системой (Northcutt, 1981). Спинной мозг птиц развит очень хорошо. Он образует большие утолщения в плечевой и поясничной области, откуда отходят нервы передних и задних конечностей (см. рис. III-12, а ). В поясничном утолщении верхняя стенка спинного мозга расходится, и центральный канал расширяется в ромбовидный синус, покрытый сверху только мозговыми оболочками. Спинномозговые нервы соединяются у птиц корешками ещё в канале позвоночного столба и выходят между дугами или через отверстия слившихся позвонков. В плечевом сплетении обычно участвуют 4 нерва, реже 3, а у страусов - только 1 нерв. Для управления задними конечностями у птиц существует два сплетения: поясничное и седалищное, или крестцовое. Поясничное сплетение обычно состоит из 3, но у страусов может включать и 5 нервов. Седалищное сплетение у всех птиц состоит из 4 крестцовых нервов. Черепно-мозговые нервы у птиц представлены 12 парами, которые начинаются по отдельности. Обонятельный нерв (I) идет от нижней поверхности обонятельной доли, которая спереди и снизу примыкает к полушариям переднего мозга. Зрительные нервы (II) после хиазмы плавно переходят в средний мозг, а блуждающий нерв, как и у рептилий, идёт из черепа далеко назад, иннервируя сердце, лёгкие, пищевод и желудок. В отличие от рептилий, добавочный нерв (XI) представлен тонкой веточкой блуждающего нерва, а подъязычный (XII) нерв имеет корешки, отходящие как от продолговатого, так и от спинного мозга (Савельев, 2001). Симпатическая нервная система птиц построена по общему для всех позвоночных принципу. Однако у птиц в шейной части расположен большой симпатический нерв, часто называемый пограничным стволом. Он лежит в канале, образованном двукорневыми началами поперечных отростков позвонков. Участок этого нерва прилежит к сонным артериям, а далее кзади симпатический нерв находится в
Рис. III-14. Строение полушария переднего мозга и зрительных долей среднего мозга птиц на примере обыкновенной неясыти (Strix aluco , а ) и сизого голубя (Columba livia , б ).
Микрофотографии соответствуют линиям, показанным на цветной схеме мозга. Оранжевый цвет - передний мозг, красный - средний мозг, фиолетовый - мозжечок, синий - задний и продолговатый мозг.Канале, образуемом головками и бугорками рёбер, и только в поясничной области он лежит свободно. По строению органов чувств птицы имеют несколько отличий от остальных животных. Осязание многих птиц (кулики, утки) сосредоточено в челюстном аппарате, где расположены специализированные механорецепторы (тельца Гранди и Хербста). Осязательные тельца расположены в коже у основания крупных перьев и сконцентрированы в восковице, покрывающей основание клюва. Совы обладают особыми осязательными перьями, расположенными вокруг клюва. У дятлов, дроздов, попугаев и глухарей осязательные функции выполняет язык, весьма богатый нервными окончаниями; в нём присутствуют преимущественно различные типы механорецепторов, а не вкусовые сосочки. Птицы пользуются языком как органом осязания. Вкусовые рецепторы у большинства птиц развиты плохо, поскольку птицы обычно заглатывают пищу, не измельчая её в клюве. Зрение птиц чрезвычайно острое. Глаз способен к аккомодации и обладает рядом морфологических особенностей, позволяющих эффективно ориентироваться, успешно охотиться и издалека определять качество пищи. В сетчатке глаза птиц плотность фоторецепторов намного больше, чем у других позвоночных. В центральной части глаза расположен наклонный гребень, где сосредоточены светочувствительные клетки. Поскольку оптическая проекция растягивается на наклонной плоскости гребня, птицы видят эту часть изображения увеличенной в 0,25-1,2 раза. Дополнительным приспособлением, улучшающим цветовосприятие, являются масляные капли в колбочках сетчатки. Они функционируют, как цветовые фильтры, что позволяет различать больше оттенков в световом диапазоне электромагнитных волн. Зрительная система птиц позволяет воспринимать объекты, излучающие в ультрафиолетовом диапазоне, и поляризованный свет. Некоторые перелётные птицы могут непосредственно воспринимать направление электромагнитных полей, что позволяет им ориентироваться в любой точке на поверхности Земли. У птиц отлично развит слух. Орган слуха состоит из внутреннего, среднего и зачатков наружного уха. Птицы воспринимают звуковые сигналы очень широкого диапазона. В этом отношении они могут намного превосходить многих млекопитающих как по диапазону, так и по чувствительности к слабым звукам. У птиц хорошо развит вестибулярный аппарат. Он состоит из полукружных каналов, отвечающих за рецепцию, связанную с угловым ускорением, и гравитационного рецептора (рецептора линейного ускорения). Все эти компоненты есть и у рептилий. Однако у птиц появился совершенный мозговой центр анализа вестибулярных и кинестетических сигналов - мозжечок.
Времени для адаптации крыльев к движениям в менее плотной среде. «Бег по воде» стал длительным биомеханическим этапом постепенного совершенствования специализированной мускулатуры и неврологических механизмов управления полётом птиц. Параллельно со становлением системы дыхания, кровообращения и мускулатуры передних конечностей у архаичных птиц возникла и теплокровность. Основной причиной её появления стала охотничья среда предков птиц. Значительное преимущество получали те животные, которые могли максимально долго пребывать в холодной воде (см. рис. III-15). Понятно, что теплокровность стала огромным достижением для птиц. Пребывая в холодной воде дольше, чем их конкуренты, они повышали вероятность добывания пищи и снижали общие энергетические затраты. С нырянием и плаванием в холодной воде, по-видимому, связан и тотальный переход предков птиц к откладке яиц. При этом способе размножения животные избавлялись от необходимости согревать и носить с собой своих будущих потомков. Это позволяло уменьшить массу тела животного, повысить плавучесть и снизить энергетические затраты при плавании и нырянии. Насиживаемые или охраняемые кладки яиц можно было увеличивать постепенно. Самки могли откладывать более крупные яйца, что привело к увеличению размеров молодых животных. Переход к плаванию и нырянию предопределил появление крыльев, развитых грудных мышц, исчезновение хвоста и развитие хвостовых желёз птиц. Перья, которые позволяли сохранять тепло при плавании и нырянии, нуждались в специальной защите от намокания. В противном случае архаичным птицам приходилось подолгу сушить своё оперение, как и некоторым современным птицам. Не исключено, что хвостовая железа появилась в эволюции птиц довольно поздно. Однако высока вероятность того, что это произошло одновременно с переходом к плаванию. Следовательно, птицы возникли в несколько этапов. На первом этапе это были небольшие оперённые рептилии, перешедшие к питанию рыбой на мелководье. В связи с новым типом питания протоптицы стали двуногими, а передние конечности долго оставались невостребованными. Обоняние было в значительной степени утрачено, а зрение стало доминирующим дистантным анализатором. На втором этапе переход к плаванию и нырянию привёл к развитию машущих передних конечностей и мощных грудных мышц. Охота в холодных водах и продолжительное ныряние стали стимулом для развития сложного дыхания, теплокровности и внеорганизменного развития эмбрионов. Передний мозг, утратив функции обонятельного анализатора,
Стал структурной базой для развития памяти, обучения и ассоциативного мышления птиц. Сложные движения в трёхмерной среде стали причиной развития мозжечка птиц как сенсомоторного центра координации движений. На третьем этапе архаичные птицы перешли к «бегу по воде» при активной работе крыльев. Такой способ быстрого перемещения над водой стал предтечей свободного полёта и создал переходные условия для медленного становления неврологических систем контроля полёта. Свободный полёт возник после увеличения размеров крыльев и изменения геометрии маховых перьев. § 45. Адаптивная радиация птиц В довольно разнообразную группу современных птиц входят летающие и нелетающие птицы. Считают, что перья, пневматичный скелет, воздушные мешки, теплокровность и другие морфологические отличия птиц сформировались в связи с адаптацией к активному полёту, поэтому бескилевые нелетающие птицы и пингвины чаще всего рассматриваются как вторично специализированные группы. Предполагается, что первоначально они были летающими, а затем утратили способность к полёту после перехода к жизни в водной среде или на земле. По-видимому, ситуация с появлением пингвинов и нелетающих птиц намного более запутанная, чем представлялось ранее. Все современные птицы имеют нервную систему чрезвычайно сходного строения. Это говорит о том, что предками птиц была одна общая группа рептилий. Они были уже настолько неврологически специализированы, что дальнейшие преобразования мозга становились модификационными. Достигнутого уровня адаптивной специализации нервной системы архаичных птиц было достаточно для успешного освоения любой доступной среды обитания. Опираясь на строение нервной системы, можно предположить следующее развитие событий. Первоначальный архетип нервной системы птиц сформировался у нелетающих рептилий при охоте на мелководье (рис. III-16, а-в ). Это событие произошло задолго до начала активного полёта. Крупный мозг, утративший обонятельные функции и специализированный по зрительно-ассоциативному типу, стал базовой структурой для появления вполне конкурентных, но не летающих животных. Вполне возможно, что часть таких архаичных птиц вернулась к жизни на поверхности земли. Они стали наземными насекомоядными или всеядными животными, которые никогда не были способны летать. Такие реверсии в эволюции птиц происходили неоднократно, но достоверно говорить о современных следах этих событий весьма затруднительно. Скорее всего единственным примером такого рода
Происхождение мозга млекопитающих
Основные признаки млекопитающих (Vertebrata ) - волосяной покров, молочные железы и теплокровность (рис. III-17). По меньшей мере два из этих признаков должны присутствовать у каждого представителя группы. У китов, моржей, броненосцев, голых землекопов и некоторых других млекопитающих волосяной покров развит незначительно или отсутствует. Основным отличием млекопитающих считают специальные железы, молоком которых самки выкармливают детёнышей. Млекопитающие имеют также сальные и потовые железы кожи. Эти железы ассоциированы с волосяным покровом и выполняют защитные, коммуникационные и репродуктивные функции. Молочные железы являются эволюционными производными специализированных кожных желёз. Только у млекопитающих есть зрелые эритроциты (красные клетки крови), лишённые ядра. У всех остальных анамний и амниот клетки крови имеют ядра. Столь же глубокие отличия млекопитающих связаны и с головным мозгом. В переднем мозге формируется шестислойная кора, а мозжечок состоит из червя и парных полушарий, которые имеют все млекопитающие (рис. III-18-III-20). Подобных образований головного мозга у других амниот нет (Савельев, 2001). § 46. Обзор классификации млекопитающих Класс современных млекопитающих разделяют на два подкласса: первозвери (Prototheria ) и настоящие звери (Theria ). К первозверям относят однопроходных млекопитающих (Monotremata ), которые могут откладывать яйца, но выкармливают детёнышей молоком и имеют своеобразный волосяной покров. Настоящие звери объединяют две обширные группы низших зверей (Metatheria ) и высших зверей (Eutheria ). Низшие звери представляют собой сумчатых млекопитающих американского и австралийского происхождения, а к высшим зверям относят всех остальных млекопитающих, объединённых в 18 обширных и часто искусственных отрядов (см. рис III-17). Основным отличием низших зверей от высших является морфофункциональный тип организации внутриматочного питания эмбрионов и плодов. Низшие звери рождают очень небольших детёнышей, которые скорее напоминают эмбрионов. Их дальнейшее развитие проходит в специальной сумке, куда открываются молочные железы. Высшие млекопитающие рождают, как правило, крупных и более сформированных детёнышей. Развитие эмбрионов и плодов высших млекопитающих обеспечивается
Документ
Менгес уже наверху! Тора распахнула двери, вышла на балкон, поежилась и взглянула на небо. А там… да, там… - там облака, подсвеченные проснувшимся солнцем, там взрыв голубого цвета – цвета кайнайта – непальского сапфира, в котором
Происхождение жизни на Земле, как устроена Вселенная, и что ждёт «человека разумного» в будущем
ДокументНаш адрес во Вселенной таков: «Вселенная, галактика Млечный Путь, Солнечная система, планета Земля». Большинство учёных считает, что Вселенная образовалась в результате Большого Взрыва, но в результате чего возник Большой Взрыв, и
Происхождение собак
ДокументПочти наверняка можно сказать, что собака была первым по времени другом человека, т.е. она была первым животным, которое ему удалось приручить. Дикарь отдаленных времен, не знавший еще ни одного домашнего животного, мог жить только
Происхождение и эволюция биовласти
ДокументВалентин ЧЕШКО, доктор философ. наук, канд. биол. наук, ст. науч. сотр. Института биологии Харьковского национального университета им. В.Н.Каразина, преподаватель кафедры философии и политологии Харьковского национального экономического
Имя материала: Основы зоологии и зоогеографии
Общая характеристика
Благодаря появлению шеи у рептилий голова подвижно сочленена с туловищем и может совершать движения не только, как у амфибий, в вертикальном направлении, но и в горизонтальном.
Туловище удлиненное (только у черепах широкое), довольно гибкое; хвост у подавляющего большинства видов длинный, способный к разнообразным движениям; ноги сильно развиты (у некоторых видов ящериц и у всех змей ноги редуцированы в связи с переходом к другому активному способу передвижения). Величина тела в среднем значительно больше, чем у амфибий, а некоторые виды достигают огромных размеров.
Покровы. Кожа покрыта роговыми прочными и в то же время легкими чешуями эктодермального происхождения, которые защищают тело не только от механических повреждений, но и от высыхания. Ороговевший слой кожи периодически сбрасывается во время линьки. У черепах и крокодилов в мезенхимном слое кожи развиваются окостенения, у остальных рептилий они отсутствуют. Черепахи хорошо защищены от врагов панцирями. Кожа современных рептилий в отличие от земноводных сухая, лишенная желез.
Окраска наружных покровов разнообразна и у многих видов имеет защитное значение, так как благодаря ей животные малозаметны на фоне окружающей среды.
Нервная система. В связи с более подвижным и сложным образом жизни нервная система рептилий развита лучше, чем у амфибий (рис. 95).
Рис. 95. Головной мозг ящерицы, вид сбоку:
1 - полушарие переднего мозга; 2 - обонятельная доля; 3 - промежуточный мозг; 4 - гипофиз; 5 - средний мозг; 6 - перекрест зрительных нервов (хиазма); 7 - мозжечок; 8 - спинной мозг
Передний мозг значительно больших размеров, чем У последних (в особенности за счет увеличения полосатых тел), на поверхности полушарий появляются нервные клетки - зачаток коры головного мозга. Поэтому у рептилий быстрее устанавлива-ются новые связи с внешней средой, чем у их предков. Возрастает роль этого отдела головного мозга как координатора жизнедеятельности всего организма. Усложняется строение промежуточного и среднего мозга, усиливается их связь с передним мозгом и другими отделами центральной нервной системы. Мозжечок в связи с усложнением способов передвижения и необходимостью сохранять равновесие тела развит заметно лучше, чем у земноводных. Увеличился и продолговатый мозг, который образует характерный для рептилий вертикальный изгиб. Спинной мозг длинный; строение его усложнилось. Это выразилось в более резком отделении наружного слоя (белого, т. е. проводящих путей) от внутреннего (серого, т. е. нервных клеток) слоя.
Из органов чувств у рептилий (за исключением змей и некоторых других видов) лучше, чем у амфибий, развиты глаза: они более подвижны. Приспособление их к видению на разных расстояниях (у многих видов на довольно больших) обеспечивается не только, как у рыб и амфибий, перемещением хрусталика, но и, как у высших позвоночных, путем изменения его кривизны. Кроме век имеется мигательная перепонка. У ряда видов есть теменной орган, связанный с промежуточным мозгом и способный к восприятию световых раздражении. Некоторые прогрессивные изменения произошли в органе слуха: немного увеличился нижний мешочек; слуховая косточка лучше передает колебания барабанной перепонки во внутреннее ухо; кроме овального окошечка появилось второе окошечко - круглое, что увеличивает подвижность экзолимфы, окружающей лабиринт; барабанная перепонка расположена не на поверхности головы, а в небольшом углублении. Органы обоняния развиты довольно хорошо, внутренняя поверхность их увеличилась. Органы вкуса у большинства видов развиты слабо. В осязании большую роль играет язык.
Скелет. Он почти полностью окостенел благодаря замене грубой волокнистой костной ткани, характерной для земноводных, тонкой волокнистой, его прочность возросла, опорная и защитная функции улучшились (рис. 96).

Рис. 96. Скелет ящерицы:
7 - ключица; 2 - лопатка; 3 - плечо; 4 - лучевая кость; 5 - локтевая кость; б - запястье; 7 - пястье; 8 - фаланги пальцев; 9 - ребра; 10 - таз; 11 - бедро; 12 - большая берцовая кость; 13 - малая берцовая кость; 14 - предплюсна; 15 - плюсна; 16 - хвостовые позвонки
Череп увеличился; вместимость мозгового отдела стала большей, чем у амфибий. Боковая поверхность черепа у древних рептилий была сплошной, позднее в ней появились височные ямы - одна или две, что уменьшило его массу и увеличило поверхность для прикрепления жевательных мышц. Нижние челюсти, как и у амфибий, подвешены к мозговому отделу черепа при помощи квадратной кости и тоже состоят из нескольких костей (зубной, сочленовной и др.).
Тело первого позвонка (атланта) приросло в виде зубца к телу второго позвонка (эпистрофея). Кольцевидный атлант получил возможность вращаться вокруг зубца эпистрофея, благодаря чему стали возможны и повороты головы, а это имело большое значение для добывания пищи (в основном других животных) и для защиты от врагов. Атлант соединен с затылочным отделом черепа с помощью одного мыщелка, что также способствует увеличению подвижности головы. Позвоночный столб разделяется на пять отделов: шейный, грудной, поясничный, крестцовый и хвостовой. Позвонки грудного отдела соединяются с грудиной хорошо развитыми ребрами, вследствие чего образуется грудная клетка, отсутствующая у амфибий. От поясничных позвонков отходят укороченные ребра, не доходящие до грудины. Крестцовых позвонков два (у амфибий один). В хвостовом отделе (за исключением черепах) позвонков много.
Кости ног лучше развиты, и они более прочные, чем у земноводных. Между костями запястья и предплюсны имеются подвижные суставы. Поэтому предплечье и проксимальный ряд запястья, а также голень и проксимальный ряд предплюсны функционируют как целостные образования. Особо нужно отметить усиление поясов конечностей: плечевого - благодаря его соединениям с грудиной и грудным отделом позвоночника, тазового - вследствие того, что он опирается на два крестцовых позвонка и обе его половины образуют два сращения - седалищное и лонное.
Мышечная система. Она подверглась дифференциации, развились новые группы мышц, от которых зависят повороты головы, движения возникшей у рептилий шеи, расширение и сужение грудной клетки и др. Значительно увеличилась мускулатура ног, развились сгибатели и разгибатели пальцев. Масса и сила всей системы возросли.
Пищеварительная система. Она более дифференцирована, чем у земноводных (рис. 97). Зубы в основном сосредоточены на челюстях, количество их велико, но они однотипны и служат преимущественно для захвата добычи. Слюнные железы лучше развиты, чем у амфибий (у ряда видов выделения этих желез ядовиты). У рептилий начинается процесс образования твердого нёба, которое особенно развито у крокодилов. Благодаря образованию твердого нёба ротовая полость отделяется от носоглоточной, что облегчает как дыхание, так и поглощение пищи. Хорошо развиты (особенно у хищников, поедающих позвоночных животных) глотка, пищевод и желудок. Печень (с желчным пузырем) и поджелудочная железа более совершенны, чем у земноводных. Тонкая кишка значительно удлинена, толстая кишка короткая и заканчивается клоакой. Современные пресмыкающиеся питаются преимущественно животными, число растительноядных видов незначительно.

Рис. 97. Внутренние органы ящерицы; А - самец; Б - самка:
/ - внешняя и 2 - внутренняя яремные вены; 3 - левая и 4 - правая сонные артерии; 5 - левая и 6 - правая дуги аорты; 7 - правое и 8 - левое предсердия; 9 - желудочек сердца; 10 - левая подключичная артерия- 11 -легкое; 12 - печень; 13 - желчный пузырь; 14 - желудок; /5 - поджелудочная железа; 16 - двенадцатиперстная кишка; 17 - толстая кишка- 18 - прямая кишка; 19 - почка; 20 - спинная аорта; 21 - выносящие вены почек; 22 - мочевой пузырь; 23 - придаток семенника; 24 - семенник; 25 - семяпровод; 26 - яичник; 27 - яйцепровод
Дыхательная система. Органами дыхания у рептилий служат легкие (см. рис. 97), так как их кожа непригодна для газообмена; слизистая оболочка ротовой полости, играющая значительную роль в дыхании земноводных, используется для выполнения этой функции немногими видами пресмыкающихся. По этой причине и особенно в связи с возросшей активностью рептилий по сравнению с амфибиями строение легких и дыхательных путей усложнилось. Легкие представляют собой органы большей величины и глубже расположены в грудной полости, чем у земноводных. Внутренняя поверхность их значительно возросла благодаря развитию на ней множества перекладин с более густой сетью кровеносных сосудов. Воздух поступает в легкие, пройдя через хоаны, глотку, гортань, длинную трахею, просвет которой поддерживается хрящевыми кольцами. Во время прохождения через трахею воздух очищается от пыли, температура его приближается к температуре тела. У амфибий трахея была в зачаточном состоянии. Задний конец трахеи разделяется на два бронха, в стенках которых тоже имеются хрящевые кольца. Осуществление вдоха и выдоха происходит более эффективно, чем у амфибий, благодаря развитию у рептилий грудной клетки, объем которой увеличивается при вдохе и уменьшается при выдохе. Такой же механизм сохранился у высших позвоночных - птиц и млекопитающих.
Кровеносная система. Сердце, как и у амфибий, трехкамерное, но в желудочке снизу появляется зачаток перегородки. У крокодилов она уже полная, и сердце у них становится четырехкамерным (два предсердия и два желудочка). Следовательно, у пресмыкающихся усиливается разделение венозного и артериального токов крови, но полной изоляции обоих токов еще нет даже у крокодилов, как это видно из нижеследующего описания (рис. 98).

Рис. 98. Сердце и основные сосуды ящерицы (светлые - сосуды с артериальной кровью, штриховка - со смешанной кровью; черные - с венозной):
1 - правое предсердие; 2 - левое предсердие; 3 - желудочек; 4 - неполная перегородка, разделяющая желудочек на левую и правую половины; 5 - легочная артерия; 6 - левая дуга аорты; 7 - правая дуга аорты; 8 - подключичная артерия; 9 - спинная аорта; 10 - общий ствол сонных артерий; 11 - левая внутренняя сонная артерия, 12 - левая наружная сонная артерия; 13 - легочная вена; 14 - задняя полая вена и передние полые (яремные) вены
В левое предсердие поступает артериальная кровь - она идет из легких. В правое предсердие поступает венозная кровь - она собирается из всех частей тела. Далее кровь из обоих предсердий поступает в желудочек: в правую его половину - венозная, в левую - артериальная. Благодаря наличию перегородки, хотя и неполной, кровь в желудочке меньше смешивается, чем у земноводных. Артериального конуса нет, и главные артериальные стволы отходят непосредственно от сердца.
От правой стороны желудочка отходят легочные артерии (бывшая четвертая пара жаберных артерий), несущие в легкие венозную кровь. Бывшая третья пара жаберных сосудов атрофировалась. Вторая пара сосудов состоит из двух дуг аорты: правая дуга начинается от левой части желудочка и несет артериальную кровь, левая дуга начинается от правой части желудочка (но ближе к левой его части) и несет смешанную кровь. От правой дуги аорты отходят сонные артерии (бывшая первая пара жаберных сосудов), идущие к голове. Они снабжают мозг, особенно чувствительный к недостатку кислорода, артериальной кровью. Обе дуги аорты соединяются в аорту, которая разветвляется на множество артерий. Таким образом, организм пресмыкающегося в основном снабжается смешанной кровью, но она насыщена большим количеством кислорода, чем у земноводных, что способствует более интенсивному обмену веществ.
Выделительная система. Она сильно изменилась в связи с усилением интенсивности обмена веществ рептилий, что привело к увеличению количества продуктов диссимиляции, и полным приспособлением их к наземному образу жизни, вследствие чего возникла необходимость более экономно использовать поступающую в организм воду. Поэтому у рептилий развились более сложные органы выделения, названные тазовыми или вторичными почками {метаяефрос). Все канальцы этих почек не имеют воронок и начинаются боуменовыми капсулами, окружающими мальпигие-вы тельца с увеличенным количеством капилляров. Канальцы тазовых почек очень длинные и густо оплетены капиллярами. В боу-меновых капсулах из просочившихся туда из мальпигиевых телец растворенных в воде продуктов диссимиляции и некоторых органических веществ (например, глюкозы) образуется первичная моча. Последняя во время прохождения через длинные почечные канальцы отдает обратно в кровь через оплетающие их стенки капилляры значительную часть воды и другие вещества, необходимые для организма. Образовавшаяся после этого вторичная моча выходит не через вольфовы каналы, а через заново развившиеся мочеточники в клоаку и накапливается в мочевом пузыре. Процессы образования первичной и вторичной мочи совершаются и в туловищных почках, но с возвратом меньшего количества воды в кровь. Первичноводные же позвоночные, в особенности земноводные, кожа которых голая, часто страдают от избытка воды, а не от ее недостатка. Следует также отметить, что у рептилий конечным продуктом разложения белков является мочевая кислота, на выведение которой из организма с мочой затрачивается мало воды. У земноводных и у ряда других первичноводных позвоночных разложение белков приводит к образованию мочевины, на удаление которой воды требуется много.
У эмбрионов рептилий есть зачаток головной почки и развитая туловищная почка, которая функционирует у взрослых особей (у ряда видов ящериц) до наступления половой зрелости. Эти факты подтверждают преемственность трех видов почек в течение эволюции позвоночных.
Размножение. В состав мужской половой системы рептилий входят два семенника округлой формы, лежащие по бокам поясничного отдела позвоночника, два придатка семенников, образовавшиеся из передних участков туловищных почек, два семяпровода (вольфовы каналы) и совокупительные органы, развившиеся из стенки клоаки. Женская половая система состоит из двух яичников, сильно разрастающихся в период размножения, и двух яйцеводов (мюллеровых каналов), впадающих в клоаку. Оплодотворение только внутреннее. Семя вводится самцами в клоаку самки при помощи совокупительных органов. Созревшие яйцеклетки, содержащие много желтка, попадают в воронки яйцеводов. Соединение женских и мужских гамет происходит в верхней части яйцеводов, куда проникают после полового акта сперматозоиды. Зиготы, продвигаясь по яйцеводам, в средней части последних окружаются белковой оболочкой, а в задней покрываются скорлупой.
У некоторых видов ящериц и змей наблюдается партеногенетическое размножение самок. Самцов у таких видов, как правило, нет.
Развитие. У рептилий развитие прямое; личинок у рептилий нет. Оплодотворенные яйца пресмыкающиеся обычно откладывают в почву, а некоторые виды - под гниющие растительные остатки. У ряда видов развитие зигот совершается в половых путях самки и сразу после выхода развившихся яиц наружу из них вылупляются сформированные животные, т. е. для таких видов характерно яйцеживорождение. У немногих видов наблюдается настоящее живорождение: сосуды желточного мешка тесно соединяются с сосудами яйцевода, и эмбрион получает значительную часть питательных веществ из организма матери. Яйцеживорождение чаще наблюдается у видов, распространенных в северных и горных районах. Очевидно, яйцеживорождение является защитой развивающихся зародышей от охлаждения и других неблагоприятных условий. Нет ни одного вида рептилий, у которого яйца развиваются в воде. Даже у рептилий, проводящих большую часть жизни в воде (все крокодилы, многие черепахи, некоторые змеи), откладка яиц и их развитие совершаются только на суше. Морские змеи, никогда не покидающие воду, относятся к живородящим.
Яйца и зародыши рептилий обладают рядом приспособлений для развития на суше. Во-первых, яйца покрыты скорлупой и другими оболочками, которые защищают их от высыхания и повреждений. Во-вторых, у зародышей развиваются особые зародышевые оболочки (серозная, амнион и аллантоис), которые имеют большое значение для обеспечения эмбрионального развития животных на суше. Серозная оболочка и амнион развиваются из одной и той же двойной складки тела зародыша, происходящей из эктодермы и мезодермы. Эта складка появляется с двух противоположных сторон в виде зачатков, которые растут навстречу друг другу и смыкаются над зародышем. Наружная часть складки превращается в серозу (ее наружный слой эктодермальный, внутренний - мезодермальный), а внутренняя - в амнион (его наружный слой мезодермальный, внутренний - эктодермальный). Полость, образуемая амнионом, наполнена водянистой жидкостью, благодаря чему развитие происходит во влажной среде, и зародыш защищен от высыхания и последствий различных сотрясений, особенно опасных в условиях воздушной среды.
Происхождение. Первые рептилии, возникшие во второй половине каменноугольного периода, обитали во влажных биотопах суши и, вероятно, размножались и развивались в воде. К этому времени суша уже была заселена низшими наземными растениями, различными беспозвоночными, начиналось завоевание воздуха насекомыми. Рептилии, которые могли удаляться на большие расстояния, чем амфибии, от водоемов, имели в изобилии пищу и не встречали серьезных врагов. В конце каменноугольного периода и неоднократно в течение пермского периода климат становился более засушливым, что вызывало гибель многих амфибий и, наоборот, содействовало прогрессивному развитию и более широкому распространению на суше рептилий. Спустя некоторое время они перешли к размножению и развитию вне водоемов, сначала в более влажных наземных биотопах, а потом в более сухих местах. В течение мезозойской эры они распространились по всему земному шару и возникло много различных групп. Наряду с хищниками "было много растительноядных видов, появились рептилии, приспособленные к полету, и рептилии, обитающие в морях. Величина их тела была различна, но большинство были крупными животными, а некоторые имели гигантские размеры - до 20 м и более в длину. Климат на протяжении почти всей мезозойской эры был теплым, очень благоприятным для этих животных, не имеющих достаточно высокой постоянной температуры тела и теряющих свою активность при охлаждении окружающей среды.
В конце мезозойской эры на земном шаре происходили мощные горообразовательные процессы, сопровождающиеся резким изменением условий существования (температуры, влажности и др.). Это повлекло за собой массовое вымирание мезозойских рептилий, не выдержавших борьбу за существование в таких тяжелых для них абиотических условиях среды, осложненную соревнованием с млекопитающими и птицами. Тогда же возникли и стали многочисленными в течение кайнозойской эры новые группы пресмыкающихся - ящерицы и произошедшие от них змеи. Из старых же групп сохранились лишь один очень примитивный вид первоящеров, небольшое количество крокодилов и черепах. Современные рептилии распространены преимущественно в теплых странах, особенно в субтропических и тропических, в странах с умеренным климатом их мало, на далеком севере они отсутствуют.
Происхождение мозга Савельев Сергей Вячеславович
§ 37. Ассоциативный центр мозга рептилий
Рассмотрев общий план строения нервной системы, следует отдельно остановиться на новых принципах организации и работы мозга, впервые реализованных у рептилий. Нервная система архаичных амниот стала логическим развитием строения удачной амфибийной конструкции. Однако мозг амфибий практически выполнял функцию сложного рефлекторного аппарата, а его интеллектуальные возможности остались невостребованными. Эволюция амфибий решалась мышцами, зубами, линейными размерами и масштабами размножения.
Шло элементарное освоение пищевых ресурсов, где для развития сложного поведения не было ни места, ни биологической необходимости. Со следами этого периода эволюции позвоночных мы сталкиваемся, пытаясь выработать условные рефлексы у различных представителей современных амфибий. Крайне низкая обучаемость и отсутствие долговременной памяти для накопления индивидуального опыта показывают, что сложных поведенческих задач перед древними амфибиями никогда в их эволюции не стояло.
Особенности развития органов чувств и признаки сложного поведения рептилий базируются на особенностях структурной организации головного мозга. Мозг рептилий отличается от мозга амфибий как в количественном, так и в качественном отношении. До появления амниот стратегии поведения или реакция на конкретный раздражитель выбирались по принципу доминантности (см. рис. III-6, е). Этот принцип состоит в том, что выраженного крупного ассоциативного центра мозга у многих первичноводных позвоночных или амфибий нет (см. рис. III-6, e). Выбор формы поведения происходит на основании сравнения активностей примерно равноценных отделов мозга, обслуживающих различные органы чувств. Решающее значение играет уровень возбуждения мозговых аналитических центров одного из анализаторов. Представительство органа чувств, достигшее в мозге наибольшего возбуждения, и становится основной областью для принятия решения. После выбора одной из инстинктивных реакций происходит её поведенческая реализация. Этот процесс осуществляется под контролем того же простого сравнения доминантностей. Если в процессе осуществления реакции возникает новое раздражение, которое изменяет соотношение возбуждений органов чувств, то поведенческая реализация инстинктивного процесса останавливается. Каждая конкретная ситуация отличается от предыдущей, но задействуется тот же набор органов чувств. Если наибольшее возбуждение достигается в той же сенсорной системе, то поведение сохраняется, а если в другой, то изменяется. Поскольку абсолютно идентичные условия в естественной жизни практически не встречаются, поведение даже самых примитивных анамний будет бесконечно разнообразно. Следовательно, поведение каждой особи будет индивидуальным с довольно высокой динамической адаптивностью.
Первые признаки ассоциативного центра появились ещё в мозге амфибий. У них таким центром мог стать средний или промежуточный мозг. Для этого были все основания. В промежуточном мозге находятся нейроэндокринные центры, контролирующие половое поведение, миграции и энергетический баланс организма анамний. Через активизацию центров промежуточного мозга запускаются инстинктивные программы поведения, которые контролируют работу других отделов мозга. Казалось бы, промежуточный мозг мог стать аналитическим центром поведения анамний, а затем и амниот. Однако в этом случае система реализации поведенческих реакций состояла бы не только в работе нервной системы. Каждый раз любое поведенческое событие приводило бы к стимуляции работы нейроэндокринных центров. Гормональная регуляция поведения реализуется долго, а неврологическая - быстро. При быстрой смене форм поведения наступал бы конфликт между инертными гормональными и динамичными нейральными программами поведения. У насекомых этот конфликт был решён в пользу нейрогормональных центров и чисто инстинктивного поведения.
С первичноводными позвоночными и амфибиями сложилась довольно нестабильная ситуация. С одной стороны, роль гормонально-инстинктивной регуляции поведения у амфибий очень велика и явно доминирует при выборе стратегий поведения. С другой стороны, нейроморфологический субстрат развит вполне достаточно для небольшой индивидуализации поведения при реализации этих стратегий. Возникла оригинальная система гормонально-доминантного выбора форм поведения из стандартного инстинктивного набора. У амфибий стратегия поведения определяется нейрогормональным состоянием особи. При реализации выбранной формы поведения она адаптируется к конкретным условиям при помощи сравнения доминантностей, которое было описано выше. В такой схеме контроля поведения анамний не остаётся места для ассоциативного центра. Он мог бы понадобиться только тогда, когда возникла бы потребность в быстрой адаптивной индивидуализации поведения. Такая ситуация может сложиться только в том случае, когда последовательная реализация инстинктивных форм поведения будет прямо зависеть от постоянно меняющейся ситуации.
В таких нестабильных условиях окружающей среды оказались архаичные рептилии. По-видимому, требования к быстрой индивидуализации поведения и памяти возросли, а реализация гормональнодоминантного принципа выбора из стандартного инстинктивного набора форм поведения стала неэффективной. Возник совершенно новый тип принятия решений, который сохранился в мозге современных рептилий, принадлежащих к отдалённым систематическим группам. Их всех объединяет одно принципиально новое качество мозга - выраженный ассоциативный центр (см. рис III-7).
Основной ассоциативный центр рептилий сформировался в крыше среднего мозга (см. рис. III-5, в; III-6; III-7, б). Он возник на базе нескольких органов чувств, которые имели представительство в этом отделе головного мозга. Основную часть крыши среднего мозга занимает представительство зрительной системы. Зрительные нервы после прохождения хиазмы перекрещиваются и поднимаются к крыше среднего мозга. Аксоны ганглиозных клеток сетчатки оканчиваются на нейронах крыши среднего мозга, которые организованы в стратифицированные структуры (см. рис. III-5, в; III-7, б). Существует чёткая топографическая связь между определённым участком сетчатки и зоной крыши среднего мозга. При этом соблюдаются форма изображения и взаимное расположение его элементов. Довольно долго переднюю часть крыши среднего мозга считали исключительно мозговым центром зрительного анализатора. Однако функционально-морфологические исследования показали, что это далеко не так.
Наряду с представительством зрительной системы в крышу среднего мозга приходят информация о соматической (кожной) чувствительности, двигательном анализаторе, вестибулярные и слуховые сигналы (см. рис. III-6, д). Слуховой анализатор у рептилий значительно увеличивает своё представительство в этом центре. В результате у многих рептилий в задней части крыши среднего мозга появляются малозаметные парные выпячивания - задние или нижние бугорки. Крыша среднего мозга становится не однородным анатомическим образованием, как у анамний, а четверохолмием. В ней сосредоточивается представительство практически всех основных дистантных и контактных анализаторов. Даже обонятельная система имеет своё представительство в крыше среднего мозга. За исключением обонятельной системы, практически все сенсорные проекции в крышу среднего мозга рептилий носят топологический характер. Это означает, что информация от каждого конкретного участка тела представлена в строго определённом участке крыши среднего мозга. Сохраняется принцип карты тела, которая точечно переносится в мозг.
Таким образом, в крыше среднего мозга рептилий сосредоточена разнообразная информация о состоянии собственного организма и окружающего мира, которая объединена по топологическому принципу. Рассмотрим, что происходит в крыше среднего мозга, если передняя правая конечность рептилии просто стоит на необычной поверхности. При оценке такой ситуации в крыше среднего мозга осуществляется сравнительный анализ соматической, сенсомоторной, слуховой и зрительной информации. Это легко сделать, поскольку все сигналы концентрируются в одном центре, а зачастую и друг над другом, как в слоистом пироге. Средний мозг проводит комплексный анализ многих факторов одного явления, что позволяет выбрать наиболее адекватную реакцию. Для этого идеально подходит стратифицированная структура крыши среднего мозга.
Предельно упрощая реальную ситуацию, можно сказать, что в крыше среднего мозга представительство различных сенсорных систем расположено на условных «этажах», организованных в горизонтальной плоскости. Каждый этаж занимает своеобразная карта. Она может быть сиюминутной информационной картой рецепторных сигналов от поверхности тела, изображением на сетчатке или акустическим полем. Все эти карты ориентированы на своих «этажах» так, что отражают примерно одно и тоже направление в пространстве. Слуховой сигнал от передней правой конечности лежит под её зрительной картой и над соматическим сигналом от кожи стопы. Специализированные «этажи» соотнесены между собой при помощи вертикальных связей, которые позволяют быстро оценить конкретную ситуацию и принять адекватное решение. Эта схема работы крыши среднего мозга позволяет понять рефлекторное быстродействие мозга рептилий. По-видимому, именно это быстродействие стало основной причиной эволюционного успеха архаичных рептилий.
Появление у рептилий совершенного рефлекторного центра принятия решений привело к нескольким важным последствиям. С одной стороны, возможность быстрого выбора решений бессмысленна, если общий уровень метаболизма будет оставаться на прежнем уровне. Следовательно, развитие среднего мозга сопровождалось повышением обмена. С другой стороны, увеличение размеров крыши среднего мозга создало необходимый клеточный субстрат для развития памяти. Индивидуальный опыт животного стал базой для сравнения событий, разнесённых во времени. Переоценить это событие трудно. Впервые возникли реальные основы для индивидуализации поведения, построенной на сравнении различных событий. Надо отметить, что эти нейробиологические преимущества мозга рептилий обычно даже не рассматриваются при реконструировании ранней эволюции амниот (Carroll, 1982).
Архаичные рептилии вслед за развитием центра рефлексологического анализа получили материальный субстрат для запоминания различных событии. Им стало доступно воспроизведение индивидуального опыта, что служит основой обучения. С самообучаемыми архаичными рептилиями амфибии уже не могли конкурировать. Гормонально-инстинктивные принципы поведения амфибий, рыб и беспозвоночных сделали их кормом для рептилий с развитым рефлекторно-ассоциативным средним мозгом.
Все перечисленные преимущества строения головного мозга рептилий не могли возникнуть сами собой. Для столь глубокой качественной перестройки мозга должны были сложиться крайне жесткие и неординарные условия. Архаичные рептилии должны были оказаться в своеобразной среде с очень высокими требованиями к аналитическим свойствам мозга и индивидуальной памяти.
Из книги Находки остатков верхнеюрских рептилий в Савельевском сланцевом руднике автора Из книги Находки остатков верхнеюрских рептилий в Савельевском сланцевом руднике автора Журавлев Константин ИвановичНаходки остатков рептилий В 1931 г., в первые же месяцы разработок сланца, когда добыча велась еще открытыми карьерами, было получено несколько находок остатков ихтиозавров. В числе этих находок серия из 13 хвостовых позвонков, найденных в пласте горючего сланца в основании
автора§ 36. Общий план строения нервной системы рептилий При масштабном освоении суши мозг рептилий изменился. Однако его морфологические перестройки, на первый взгляд, не были столь принципиальными, как у амфибий. Поверхностный анатомический анализ показывает, что древний
Из книги Происхождение мозга автора Савельев Сергей Вячеславович§ 38. Условия возникновения мозга рептилий Ассоциативный мозговой центр не может возникнуть случайно. Энергетические затраты на содержание ассоциативного центра мозга и цена смены стратегий поведения всегда очень высоки (см. главу I). Должна быть причина для подобных
Из книги Происхождение мозга автора Савельев Сергей Вячеславович§ 40. Адаптивная радиация архаичных рептилий Небольшие хищные и всеядные рептилии, эволюционировавшие в агрессивной среде растительных завалов, не были самыми крупными животными того времени. В конце карбона размер их тела обычно не превышал нескольких десятков
Из книги Биологическая химия автора Лелевич Владимир ВалерьяновичАктивный центр фермента Участок молекулы фермента, который специфически взаимодействует с субстратом, называется активным центром. Активный центр – это уникальная комбинация аминокислотных остатков в молекуле фермента, обеспечивающая непосредственное
автораГде находится центр масс системы Земля – Луна? Центр масс системы Земля – Луна, так называемый барицентр, находится на расстоянии 4672 километра от центра Земли по направлению к Луне, то есть на глубине приблизительно 1700 километров под поверхностью Земли. Строго говоря, по
Из книги Новейшая книга фактов. Том 1. Астрономия и астрофизика. География и другие науки о Земле. Биология и медицина автора Кондрашов Анатолий ПавловичЧто такое центр удовольствия и где он расположен в организме? Одной из частей головного мозга является гипоталамус, являющийся отделом промежуточного мозга и расположенный под зрительными буграми (таламусом). Гипоталамус, в котором расположены центры вегетативной
автора Кондрашов Анатолий Павлович Из книги Новейшая книга фактов. Том 1 [Астрономия и астрофизика. География и другие науки о Земле. Биология и медицина] автора Кондрашов Анатолий Павлович Из книги Фармацевтическая и продовольственная мафия автора Броуэр ЛуиЦентр фармакологического надзора «Париж, 4 февраля 1977 г. НЦФН, созданный в январе 1974 г., 2 февраля собрался на Генеральную ассамблею вместе с делегатами Национальных хартий врачей и фармацевтов, к которым присоединилась группа центров по борьбе с интоксикацией и
Из книги В дебрях времени автора Чижевский Герман МихайловичКлассификация и краткая характеристика основных групп амфибий, рептилий, птиц и млекопитающих КЛАСС АМФИБИИ, ИЛИ ЗЕМНОВОДНЫЕПодкласс апсидоспондилыНадотряд ЛабиринтодонтыГруппа амфибий, вымершая не позже триасового периода и достигшая расцвета в конце палеозойской
автора Кандель Эрик Ричард Из книги В поисках памяти [Возникновение новой науки о человеческой психике] автора Кандель Эрик Ричард Из книги Психопаты. Достоверный рассказ о людях без жалости, без совести, без раскаяния автора Кил Кент А.Мендотский реабилитационный центр для несовершеннолетних В начале 1990-х США захлестнула настоящая эпидемия подросткового насилия. Число преступлений, совершаемых несовершеннолетними, почти удвоилось между 1980 и 1993 годами. Казалось, ничто не может остановить этот
Из книги Читая между строк ДНК [Второй код нашей жизни, или Книга, которую нужно прочитать всем] автора Шпорк ПетерИз Берлина в центр будущей революции Он не обижается, когда его принимают за студента или докторанта. А это происходит с 32-летним генетиком постоянно. Александр Майсснер - серо-голубые глаза, темно-русые небрежно причесанные волосы, трехдневная щетина - не только молод,
Анатомия, морфология и экология пресмыкающихся
10. Нервная система, органы чувств и нервная деятельность пресмыкающихся
Головной мозг пресмыкающихся отличается от головного мозга земноводных рядом важных особенностей и нередко выделяется в особый «зауропсидный тип », присущий также птицам и противопоставляемый «ихтиопсидному типу» мозга земноводных и рыб (рис. 16).

Рис. 16. Схема организации головного мозга пресмыкающихся (по Никитенко, 1969): 1 - обонятельная луковица, 2 - обонятельный тракт, 3 - базальное ядро и гипопаллиум, 4 - стриатум, б- неопаллиум, 6,7 - дорзальная и вентральная части зрительного бугра, 8 - зрительный тракт, 9 - эпифиз, 10 - зрительная кора среднего мозга, 11 и 12 - ретикулярная формация среднего и продолговатого мозга, 13 - слуховое ядро среднего мозга, 14 - продолговатый мозг, 15 - мозжечок, 16 - чувствительные ядра продолговатого мозга, 17 - чувствительные пучки волокон кожно-мышечного анализатора, 18 -- двигательные волокна к мышцам тела, 19 - то же, к мышцам головы, 20 - гипофиз
Передний мозг рептилий более крупен; его увеличение связано с развитием мозгового свода полушарий и заметному увеличению лежащих на дне полосатых тел (corpora striata), составляющих большую часть массы переднего мозга (рис. 17).

Рис. 17. (по Паркеру). А - сверху; Б - снизу; В - сбоку: 1 - передний мозг, 2 - полосатое тело, 3 - средний мозг, 4 - мозжечок, 5 - продолговатый мозг, 6 - воронка, 7 - гипофиз, 8 - хиазма, 9 - обонятельные доли, 10 - эпифиз, II-XII - головные нервы
В мозговом своде полушарий отчетливо различаются первичный свод , или архипаллиум , занимающий большую часть крыши полушарий, а также зачаток неопаллиума . У современных рептилий роль основного ассоциативного центра, определяющего характер поведения, играют передний мозг и кора среднего мозга, связанные через центры промежуточного мозга. При этом первичная функция переднего мозга - обработка обонятельной информации - сохраняется, но становится второстепенной; с обонятельными луковицами полушария соединяются длинным обонятельным трактом.
Обонятельный центр по сравнению с амфибиями более сложен и дифференцирован. В переднем мозге рептилий возникают экранные структуры , обычно свидетельствующие об усложненной нервной деятельности. <Экранными структурами называют послойное расположение тел нейронов, их аксонов и дендритов, свидетельствующее о высокой упорядоченности мозговых структур и обеспечивающее обработку информации, получаемой от рецепторов. Экранные структуры особо характерны для новой коры мозга млекопитающих.>
Промежуточный мозг сверху прикрыт полушариями переднего мозга. В его крыше расположены эпифиз (эндокринная железа) и теменной орган , у гаттерии и некоторых ящериц служащий дополнительным фоторецептором : он способен воспринимать сигналы опасности (быстрое затенение) и, видимо, служит рецептором, регистрирующим сезонные изменения светового режима. Уплотненный и прозрачный передний отдел теменного органа напоминает хрусталик глаза, а его бокаловидная задняя часть снабжена пигментными и чувствующими клетками. Дно промежуточного мозга участвует в работе эндокринной системы в качестве нейросекреторной доли гипофиза, связанной с гипоталамической областью промежуточного мозга. Этим путем гипофиз получает информацию о состоянии внешней среды, собранную органами чувств и обработанную мозговыми центрами, что позволяет ему координировать работу всей эндокринной системы. Средствами связи служат воротные системы кровеносных сосудов, существующие во всех долях гипофиза. Они, видимо, получают и передают информацию, кодированную в химических соединениях.
Зрительная кора среднего мозга более развита, нежели у амфибий. Сохраняя значение основного центра обработки зрительной информации, у рептилий она принимает участие и в формировании актов сложного поведения . В отличие от амфибий мозжечок пресмыкающихся крупный, что отвечает большей сложности и интенсивности их движений.
Продолговатый мозг образует изгиб в вертикальной плоскости, характерный для всех амниот. Сохраняя значение центра автоматической (безусловнорефлекторной) двигательной активности и основных вегетативных функций (дыхания, кровообращения, пищеварения и др.), продолговатый мозг находится под большим контролем передних отделов мозга. В связи с этим его значение как центра ассоциативной деятельности уменьшается. Имеются 11 пар головных нервов.
В спинном мозге отчетливее выражено разделение белого (проводящих путей) и серого (нервных клеток) вещества. Это свидетельствует об усилении контроля центров головного мозга над рефлекторными механизмами спинного мозга. В области плечевого и тазового поясов спинной мозг образует нервные сплетения, обеспечивающие обслуживание системы мускулов конечностей. Вегетативная (симпатическая и парасимпатическая) нервная система в виде цепи парных нервных ганглиев, связанных со спинно-мозговыми корешками и между собой промежуточными комиссурами, выражена отчетливо и представляет механизм, координирующий работу вегетативных систем организма в условиях изменчивой среды. Нельзя не видеть в этом связи с общей интенсификацией двигательной активности пресмыкающихся. С усложнением движений и развитием общения между животными связано прогрессивное развитие органов чувств.
Орган зрения приспособлен к работе в воздушной среде. Глаз защищен наружными веками и мигательной перепонкой. У змей и некоторых ящериц (гекконов , сцинков , части безногих ящериц) веки срастаются, образуя прозрачную оболочку . У ночных видов глаза обычно увеличены и имеют вертикальный зрачок . Слезные железы предохраняют глаз от высыхания, увлажняя поверхность глазного яблока. В отличие от лягушек глазные яблоки не могут втягиваться в ротовую полость и совершают только вращательные движения. У хамелеонов каждый глаз может двигаться самостоятельно, что важно при подкарауливании добычи, когда тело неподвижно. В меньшей степени способностью к раздельному движению глаз наделены агамы (p. Calotes) и некоторые игуаны.
Аккомодация глаза достигается перемещением хрусталика и изменением его кривизны с помощью поперечнополосатого ресничного мускула. От задней стенки глазного яблока в стекловидное тело вдается «гребень» - богатый кровеносными сосудами пигментированный вырост; видимо, он улучшает питание сетчатки. Гребень лучше развит у обитателей открытых пространств. Сетчатка глаза пресмыкающихся сложнее, чем сетчатка земноводных. У части ночных видов она содержит только палочки. У обладающих цветным зрением дневных видов в сетчатке есть и палочки, и колбочки; у многих видов колбочки снабжены своеобразными светофильтрами в виде бесцветных или окрашенных (желтых, оранжевых, красных) жировых капель. Чувствительность цветового зрения большинства пресмыкающихся смещена в желто-оранжевую часть спектра. В отличие от земноводных анализ и синтез зрительных восприятий производится не в сетчатке, а преимущественно в зрительной коре среднего мозга.
В пространственной ориентации и общении пресмыкающихся зрение играет важную, обычно решающую роль. С этим связано существование яркой, иногда сложной демонстративной окраски , имеющей опознавательное значение. Ту же роль выполняют позы . Многие детали окраски и ряд сигнальных движений служат защитой от врагов и конкурентов: устрашение раскрыванием воротника австралийской плащеносной ящерицей или разинутой пастью ушастой круглоголовки , окрашенной приливающей кровью в ярко-красный цвет, и т.п. Взмахи хвоста песчаной круглоголовки , раскрашенного черно-белыми полосами, служат сигналом прекращения опасности.
У ямкоголовых змей (Crotalidae), у питонов (Pythoninae) и африканских гадюк (Bitis) имеются особые органы термического чувства - терморецепторы и даже термолокаторы . Термолокаторы ямкоголовых змей - парные ямки, расположенные по бокам морды, между ноздрями и глазами; у питонов подобные же неглубокие ямки имеются на верхнегубных щитках, а у африканских гадюк чашеобразные углубления лежат позади ноздрей.
Лучше изученный термолокатор ямкоголовых змей состоит из прикрытой прозрачной пленкой ямки и меньших размеров внутренней полости, разделенных тонкой (15 мкм) мембраной; иннервируется ветвью тройничного нерва. Внутренняя полость сообщается с внешней средой тонким каналом, запираемым кольцевым мускулом. При закрытом канале излучаемое жертвой тепло (поток инфракрасных лучей), нагревая наружную полость, увеличивает давление на перепонку и по разности давления в правой и левой полостях позволяет определить направление источника излучения, т. е. лоцировать добычу в темноте (например, грызуна в его норе). Полагают, что эти терморецепторы способны регистрировать изменения температуры в тысячные доли градусов .
Орган слуха пресмыкающихся в схеме близок к органу слуха лягушек. Он состоит из среднего уха с барабанной перепонкой и одной слуховой косточкой - стремечком , передающим колебания перепонки на круглое окошко, отделяющее полость внутреннего уха. Этот механизм усиливает звуки , распространяющиеся в воздушной среде. Во внутреннем ухе обособляется улитка (lagena) , служащая аппаратом анализа и кодирования акустических сигналов. Улитка еще не сложна и у большинства видов представляет мешкообразный вырост. Это соответствует относительно небольшой роли слуха в жизни пресмыкающихся. Они воспринимают звуки в диапазоне 20-6000 герц (Гц), но большинство хорошо слышит только в диапазоне 60-200 Гц. Крокодилы лучше воспринимают звуки частотой 100-3000 Гц. У змей слух особенно слаб; они лишены барабанной перепонки и воспринимают преимущественно звуки, распространяющиеся по субстрату (земле) или в воде (так называемый сейсмический слух ). То же свойственно и змеевидным ящерицам. Передача звуков с субстрата на овальное окошко среднего уха обеспечивается квадратной и квадратно-скуловой костями. Сравнительно ограниченный слух змей соответствует их слабым голосовым возможностям. Невысоки и слуховые способности черепах , барабанная перепонка которых толстая, а слуховой проход у некоторых видов закрыт утолщенной кожей.
Большинство пресмыкающихся немо ; их звуковой мир беден. Звуки змей (шипенье, хрип, стук хвостовых погремушек) и некоторых ящериц (скрип чешуи) чаще служат угрожающим предупреждением . Громкие ревущие звуки издают крокодилы; они используются при охране территории и при поисках особи другого пола.
Хеморецепторы играют в ориентации и общении рептилий важную, хотя и меньшую по сравнению со зрением роль. Органы обоняния открываются наружу парными ноздрями , а в полость рта - щелевидными хоанами . Срединная часть носоглоточного хода дифференцирована на верхний обонятельный и нижний дыхательный отделы. Обонятельный отдел имеет складчатые стенки, увеличивающие его поверхность. Впереди хоан в крыше ротовой полости имеется углубление, так называемый якобсонов орган . Он воспринимает запах находящейся во рту пищи или веществ, которые животное собирает с земли своим подвижным языком и вносит в ротовую полость.
Чувствительность обоняния пресмыкающихся выше, чем у земноводных. Многие ящерицы с помощью обоняния находят пищу, выкапывая ее из песка с глубины до 6-8 см. Вараны , полозы и гадюки с его помощью находят пищу, разыскивают особей другого пола и могут различать особей своего или чужих видов; реагируют они и на запахи врагов. Черепахи , ящерицы и крокодилы имеют специальные пахучие железы , секретом которых они метят занятую территорию, предупреждая вторжение на нее чужих особей.
Чувство осязания выражено отчетливо. Черепахи ощущают даже легкое прикосновение к панцирю. У многих ящериц имеются осязательные «волоски» , образовавшиеся из ороговевших клеток кожи и расположенные по краям чешуи.
В экологическом Центре "Экосистема" можно приобрести